Inflamación neurogénica, ou NI, é o proceso fisiolóxico onde os mediadores son descargados directamente dos nervios cutáneos para comezar unha resposta inflamatoria. Isto orixina a creación de reaccións inflamatorias locais, incluíndo eritema, inchazo, aumento da temperatura, tenrura e dor. As fibras C-somáticas aferentes finas non amelinadas, que responden a estímulos mecánicos e químicos de baixa intensidade, son en gran parte responsables da liberación destes mediadores inflamatorios.
Cando se estimulan, estas vías nerviosas nos nervios cutáneos liberan neuropéptidos enerxéticos ou péptido relacionado co xene calcitonina (CGRP) rapidamente no microambiente, provocando unha serie de respostas inflamatorias. Hai unha distinción significativa na inflamación inmunogénica, esa é a primeira resposta protectora e reparadora do sistema inmune cando un patóxeno entra no corpo, mentres que a inflamación neurogénica implica unha conexión directa entre o sistema nervioso e as respostas inflamatorias. Aínda que a inflamación neurogénica ea inflamación inmunolóxica poden existir ao mesmo tempo, os dous non son clásicamente indistinguibles. O seguinte artigo é discutir o mecanismo da inflamación neurogénica e o papel do sistema nervioso periférico na defensa e inmunopatoloxía do hóspede.
Contidos
Inflamación neuroxénica: o papel do sistema nervioso periférico na defensa do hóspede e na inmunopatoloxía
Abstracto
Os sistemas nerviosos e inmunitarios periféricos son tradicionalmente pensados para servir funcións separadas. Esta liña está, porén, cada vez máis borrosa por novos coñecementos sobre a inflamación neurogénica. As neuronas nociceptoras posúen moitas das mesmas vías de recoñecemento molecular para o perigo como células inmunes e en resposta ao perigo, o sistema nervioso periférico comunícase directamente co sistema inmunitario, formando un mecanismo de protección integrado. A densa rede de inervación das fibras sensoriais e autonómicas nos tecidos periféricos ea alta velocidade da transducción neural permite unha rápida modificación neurogénica local e sistémica da inmunidade. As neuronas periféricas tamén parecen desempeñar un papel importante na disfunción inmune nas enfermidades autoinmunes e alérxicas. Polo tanto, a comprensión da interacción coordinada das neuronas periféricas con células inmunes pode avanzar en enfoques terapéuticos para aumentar a defensa do hóspede e suprimir a inmunopatoloxía.
introdución
Hai dous mil anos, Celsus definiu a inflamación como a participación de catro signos cardinais: Dolor (dor), Calor (calor), Rubor (vermelhidão) e Tumor (inchazo), unha observación que indica que a activación do sistema nervioso foi recoñecida como parte integral de inflamación. Non obstante, a dor pensouse principalmente desde entón, só como un síntoma e non como un participante na xeración de inflamación. Nesta perspectiva, demostramos que o sistema nervioso periférico xoga un papel directo e activo na modulación da inmunidade innata e adaptativa, de xeito que o sistema inmune e nervioso pode ter unha función protectora integrada común na defensa do hóspede e na resposta á lesión dos tecidos, un intrincado interacción que tamén pode levar a patoloxía en enfermidades alérxicas e autoinmunes.
A supervivencia dos organismos depende fundamentalmente da capacidade de defensa contra o dano potencial causado por danos e infeccións nos tecidos. A defensa do hóspede implica un comportamento de evitación para eliminar o contacto cun ambiente perigoso (nocivo) (unha función neuronal) e unha neutralización activa de axentes patóxenos (unha función inmune). Tradicionalmente, o papel do sistema inmune na loita contra os axentes infecciosos e na reparación de lesións nos tecidos considerouse bastante distinto do do sistema nervioso, que transdúe sinais ambientais e internos danantes en actividade eléctrica para producir sensacións e reflexos (Fig. 1). Propoñemos que estes dous sistemas son realmente compoñentes dun mecanismo de defensa unificado. O sistema nervioso somatosensorial está ideal para detectar o perigo. En primeiro lugar, todos os tecidos altamente expostos ao ambiente externo, como as superficies epiteliais da pel, os pulmóns, o tracto urinario e dixestivo, están densamente inervados por nociceptores, fibras sensoriais que producen dor de alto limiar. En segundo lugar, a transducción de estímulos externos nocivos é case instantánea, as ordes de magnitude máis rápidas que a mobilización do sistema inmune innato e, polo tanto, poden ser o "primeiro respondedor" na defensa do hóspede.
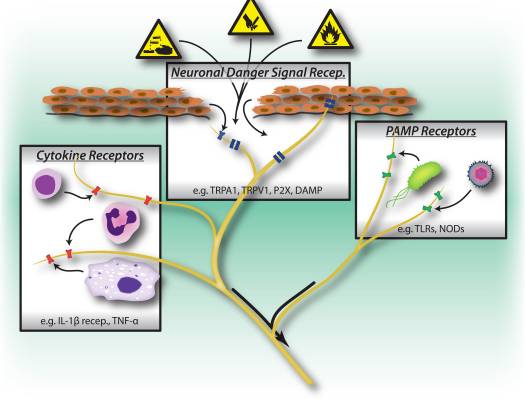
Imaxe 1: Os estímulos nocivos, as vías de recoñecemento microbianas e inflamatorias accionan o sistema nervioso periférico. As neuronas sensoriais teñen varios medios para detectar a presenza de estímulos nocivos / nocivos. 1) Os receptores de sinal de perigo, incluídos os canles TRP, os canles P2X e os patróns moleculares asociados ao perigo (DAMP) recoñecen sinais exóxenos do medio (por exemplo, calor, acidez, produtos químicos) ou sinais endóxenos de perigo liberados durante o trauma / lesión tisular (por exemplo, ATP, ácido úrico, hidroxinonales). 2) Os receptores de recoñecemento de patrón (PRR) como os receptores tipo Toll (TLR) e os receptores tipo Nod (NLR) recoñecen os patóxenos asociados aos patóxenos (PAMPs) arroxados pola invasión de bacterias ou virus durante a infección. 3) Os receptores de citocinas recoñecen factores secretados por células inmunitarias (por exemplo, IL-1beta, TNF-alfa, NGF), que activan mapas e outros mecanismos de sinalización para aumentar a excitabilidade da membrana.
Ademais das entradas ortodrómicas á medula espiñal e ao cerebro desde a periferia, os potenciais de acción nas neuronas nociceptoras tamén se poden transmitir antidrómicamente nos puntos de ramificación cara á periferia, o reflexo do axón. Estes xunto con despolarizacións locais sostidas levan a unha liberación rápida e local de mediadores neuronais tanto dos axóns periféricos como dos terminais (Fig. 2) 1. Os experimentos clásicos de Goltz (en 1874) e de Bayliss (en 1901) demostraron que as raíces dorsais estimulantes electricamente induce vasodilatación da pel, o que levou ao concepto de "inflamación neuroxénica", independentemente da producida polo sistema inmunitario (Fig. 3).
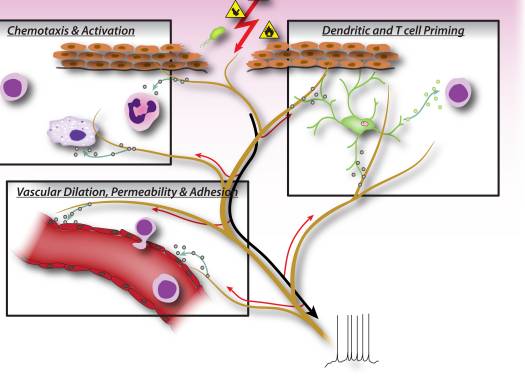
Imaxe 2: Os factores neuronales liberados das neuronas sensoriais do nociceptor levan directamente a quimioxia do leucocito, a hemodinámica vascular ea resposta inmune. Cando os estímulos nocivos activan sinais aferentes nos nervios sensoriais, xéranse reflexos do axón antidromático que inducen a liberación de neuropéptidos nos terminales periféricos das neuronas. Estes mediadores moleculares teñen varias accións inflamatorias: 1). Quimiotáxica e activación de neutrófilos, macrófagos e linfocitos no lugar da lesión e desgranulación de mastocitos. 2) Sinalización a células endoteliais vasculares para aumentar o fluxo sanguíneo, fuga vascular e edema. Isto tamén permite un reclutamento máis fácil dos leucocitos inflamatorios. 3) Priming das células dendríticas para dirixir a diferenciación posterior de células T helper en subtipos Th2 ou Th17.

Imaxe 3: Cronoloxía dos avances na comprensión dos aspectos neuroxénicos da inflamación de Celsus ata a actualidade.
A inflamación neuroxénica está mediada pola liberación dos neuropéptidos, péptido relacionado co xene da calcitonina (CGRP) e substancia P (SP) dos nociceptores, que actúan directamente sobre as células do endotelio vascular e do músculo liso 2. O CGRP produce efectos de vasodilatación 5, 2, mentres que SP aumenta a permeabilidade capilar levando á extravasación do plasma e edema 3, 4, contribuíndo ao rubor, calor e tumor de Celsus. Non obstante, os nociceptores liberan moitos neuropéptidos adicionais (base de datos en liña: www.neuropeptides.nl/), incluíndo adrenomedullina, neurocininas A e B, péptido intestinal vasoactivo (VIP), neuropéptido (NPY) e péptido liberador de gastrina (GRP), así como outros mediadores moleculares como glutamato, óxido nítrico (NO) e citocinas como a eotaxina 6.
Agora apreciamos que os mediadores liberados das neuronas sensoriais na periferia non só actúan sobre a vasculatura, senón que tamén atraen e activan directamente as células inmunes innatas (mastocitos, células dendríticas) e as células inmunes adaptativas (linfocitos T) 7-12. No contexto agudo do dano nos tecidos, conxecturamos que a inflamación neuroxénica é protectora, facilitando a curación fisiolóxica das feridas e a defensa inmune contra os patóxenos activando e recrutando células inmunes. Non obstante, estas comunicacións neuroinmunes tamén desempeñan un papel importante na fisiopatoloxía das enfermidades alérxicas e autoinmunes amplificando as respostas inmunes patolóxicas ou inadaptadas. En modelos animais de artrite reumatoide, por exemplo, Levine e colegas demostraron que a denervación da articulación leva a unha atenuación sorprendente na inflamación, que depende da expresión neuronal da substancia P 13, 14. En estudos recentes sobre a inflamación alérxica das vías respiratorias, colite e psoríase, as neuronas sensoriais primarias xogan un papel central na iniciación e aumento da activación da inmunidade innata e adaptativa 15-17.
Propuxemos, polo tanto, que o sistema nervioso periférico non só desempeña un papel pasivo na defensa do hóspede (detección de estímulos nocivos e iniciación do comportamento de evitación), senón tamén un papel activo en conxunto co sistema inmunitario na modulación das respostas e o combate a efectos nocivos. estímulos, un papel que pode ser subvertido para contribuír á enfermidade.
Camiños de recoñecemento de perigos compartidos nos sistemas inmunes nerviosos e innatos periféricos
As neuronas sensoriais periféricas están adaptadas para recoñecer o perigo ao organismo en virtude da súa sensibilidade aos intensos estímulos químicos, térmicos e irritantes (Fig. 1). Os canais iónicos do potencial de receptor transitorio (TRP) son os mediadores moleculares máis ampliamente estudados da nocicepción, realizando unha entrada non selectiva de cationes despois da activación por varios estímulos nocivos. O TRPV1 é activado por altas temperaturas, baixo pH e capsaicina, o compoñente irritante vallinoide dos chiles 18. TRPA1 media a detección de produtos químicos reactivos, incluídos irritantes ambientais como o gas lacrimóxeno e os isotiocianatos industriais 19, pero o máis importante é que tamén se activa durante a lesión tisular por sinais moleculares endóxenos, incluíndo 4-hidroxinonal e prostaglandinas 20, 21.
Curiosamente, as neuronas sensoriais comparten moitas das mesmas rutas de receptores de recoñecemento molecular de patóxeno e perigo como células inmunidades innatas, que lles permiten tamén detectar patóxenos (Fig. 1). No sistema inmunitario, os patógenos microbianos detectáronse os receptores de recoñecemento de patróns codificados en xermina (PRR), que recoñecen patóxenos moleculares asociados aos patóxenos exóxenos (PAMP). Os primeiros PRRs que se identificaron eran membros da familia do receptor de peaxe (TLR), que se unían a léveda, compoñentes de parede celular derivada de bacterias e ARN viral 22. Despois da activación do PRR, encóntranse as vías de sinalización descendente que inducen a produción de citocinas e activan a inmunidade adaptativa. Ademais dos TLR, as células inmunitarias innatas activáronse durante a lesión tisular por sinais de perigo derivados endóxenos, tamén coñecidos como patróns moleculares asociados ao dano (DAMP) ou alarmins 23, 24. Estes sinais de perigo inclúen HMGB1, ácido úrico e proteínas de choque térmico liberadas por morrer células durante a necrose, activando células inmunes durante respostas inflamatorias non infecciosas.
Os PRR incluídos os TLR 3, 4, 7 e 9 exprésanse por neuronas nociceptoras e a estimulación por ligandos TLR leva á indución de correntes internas e á sensibilización dos nociceptores a outros estímulos de dor 25-27. Ademais, a activación de neuronas sensoriais polo ligando TLR7 imiquimod leva á activación dunha vía sensorial específica de coceira 25. Estes resultados indican que a dor e a picazón asociadas á infección poden deberse en parte á activación directa das neuronas por factores derivados do patóxeno, que á súa vez activan as células inmunes a través da liberación periférica de moléculas de sinalización neuronal.
Un DAMP / alarma importante liberado durante a lesión celular é o ATP, que é recoñecido polos receptores purinérxicos tanto nas neuronas nociceptoras como nas células inmunes 28-30. Os receptores purinérxicos están formados por dúas familias: receptores P2X, canles de catión con gandos e receptores P2Y, receptores acoplados á proteína G. Nas neuronas nociceptoras, o recoñecemento do ATP prodúcese a través de P2X3, o que leva a correntes de catións densensibilizándose rapidamente e a dor 28, 30 (Fig. 1), mentres que os receptores P2Y contribúen á activación do nociceptor sensibilizando TRP e as canles de sodio con tensión. Nos macrófagos, a unión do ATP aos receptores P2X7 leva á hiperpolarización e á activación augas abaixo do inflammasoma, un complexo molecular importante na xeración de IL-1beta e IL-18 29. Polo tanto, o ATP é un sinal de perigo potente que activa tanto as neuronas periféricas como as innatas. inmunidade durante a lesión, e algunhas evidencias suxiren que as neuronas expresan partes da maquinaria molecular inflammasoma 31.
O flip side dos sinais de perigo nos nociceptores é o papel das canles TRP na activación da célula inmune. O TRPV2, un homólogo do TRPV1 activado por calor nocivo, está expresado a altos niveis de células inmunitarias innatas 32. A ablación xenética de TRPV2 provocou defectos na fagocitosis macrófago e eliminación de infeccións bacterianas 32. Os mastocitos tamén expresan canles TRPV, que poden mediar directamente a súa degranulación 33. Queda por determinar se as sinais endóxenos de perigo activan as células inmunitarias dun xeito similar aos nociceptores.
Un medio clave de comunicación entre as células inmunes e as neuronas nociceptoras é a través das citocinas. Tras a activación dos receptores de citoquinas, as vías de transdución de sinais actívanse nas neuronas sensoriais que levan á fosforilación augas abaixo das proteínas da membrana, incluíndo TRP e canles con tensión (Fig. 1). A sensibilización resultante dos nociceptores significa que agora os estímulos mecánicos e de calor normalmente inocuos poden activar os nociceptores. A interleucina 1 beta e o TNF-alfa son dúas citocinas importantes liberadas polas células inmunes innatas durante a inflamación. IL-1beta e TNF-alfa son detectados directamente por nociceptores que expresan os receptores cognitivos, inducindo a activación das pinas do mapa p38 conducindo a un aumento da excitabilidade da membrana 34-36. O factor de crecemento nervioso (NGF) e a prostaglandina E (2) tamén son os principais mediadores inflamatorios liberados das células inmunes que actúan directamente sobre as neuronas sensoriais periféricas para causar sensibilización. Un efecto importante da sensibilización nociceptora por factores inmunes é un aumento da liberación de neuropéptidos nos terminais periféricos que activan aínda máis as células inmunes, inducindo así un bucle de retroalimentación positiva que impulsa e facilita a inflamación.
Control do sistema nervioso sensorial de inmunidade innata e adaptativa
Nas fases iniciais da inflamación, as neuronas sensoriais sinalizan aos mastocitos e ás células dendríticas residentes nos tecidos, que son células inmunes innatas importantes para iniciar a resposta inmune (Fig. 2). Os estudos anatómicos demostraron unha aposición directa de terminais con mastocitos, así como con células dendríticas, e os neuropéptidos liberados dos nociceptores poden inducir a degranulación ou a produción de citocinas nestas células 7, 9, 37. Esta interacción xoga un papel importante nas vías respiratorias alérxicas. inflamación e dermatite 10-12.
Durante a fase efectora da inflamación, as células inmunes necesitan atopar o camiño para o lugar específico da lesión. Moitos mediadores liberados de neuronas sensoriais, neuropéptidos, quimiocinas e glutamato son quimiotácticos para neutrófilos, eosinófilos, macrófagos e células T e melloran a adhesión endotelial que facilita a inserción das células inmunes 6, 38-41 (Fig. 2). Ademais, algunhas evidencias implican que as neuronas poden participar directamente na fase efectora, xa que os propios neuropéptidos poden ter funcións antimicrobianas directas 42.
As moléculas de sinalización derivadas neuronalmente tamén poden dirixir o tipo de inflamación, contribuíndo á diferenciación ou especificación de diferentes tipos de células T inmunes adaptativas. Un antíxeno fagocítase e é procesado por células inmunes innatas, que despois migran ao ganglio linfático máis próximo e presentan o péptido antigénico ás células T. Dependendo do tipo de antíxeno, as moléculas costimuladoras da célula inmune innata e as combinacións de citocinas específicas, as células T nave maduran en subtipos específicos que mellor serven o esforzo inflamatorio para limpar o estímulo patóxeno. As células T CD4, ou células T auxiliares (Th), pódense dividir en catro grupos principais, as células reguladoras Th1, Th2, Th17 e T (Treg). As células Th1 están principalmente implicadas na regulación das respostas inmunes a microorganismos intracelulares e enfermidades autoinmunes específicas de órganos; O Th2 é fundamental para a inmunidade contra os axentes patóxenos extracelulares, como os helmintos, e son responsables das enfermidades inflamatorias alérxicas; As células Th17 xogan un papel central na protección contra desafíos microbianos, como bacterias e fungos extracelulares; As células Treg están implicadas no mantemento da autotolerancia e na regulación das respostas inmunes. Este proceso de maduración das células T parece estar moi influído polos mediadores neuronais sensoriais. Os neuropéptidos, como CGRP e VIP, poden polarizar as células dendríticas cara a unha inmunidade de tipo Th2 e reducir a inmunidade de tipo Th1 promovendo a produción de certas citocinas e inhibindo outras, así como reducindo ou mellorando a migración de células dendríticas aos ganglios linfáticos locais 8 , 10, 43. As neuronas sensoriais tamén contribúen considerablemente á inflamación alérxica (principalmente Th2) 17. Ademais de regular as células Th1 e Th2, outros neuropéptidos, como SP e Hemocinina-1, poden impulsar a resposta inflamatoria máis cara a Th17 ou Treg 44, 45, o que significa que as neuronas tamén poden estar implicadas na regulación da resolución inflamatoria. En inmunopatoloxías como a colite e a psoríase, o bloqueo de mediadores neuronais como a substancia P pode amortecer significativamente as células T e o dano mediado por inmunidade 15-17, aínda que antagonizar un mediador só pode ter un efecto limitado sobre a inflamación neuroxénica.
Tendo en conta que as moléculas de sinalización liberadas das fibras nerviosas sensoriais periféricas regulan non só pequenos vasos sanguíneos, senón tamén a quimiotaxis, a maduración e a activación das células inmunitarias, está quedando claro que as interaccións neuro-inmunes son moito máis complexas do que se pensaba anteriormente (Fig. 2). Ademais, é bastante concebible que non sexan mediadores neuronales individuais senón combinacións específicas de moléculas de sinalización liberadas de nociceptores que inflúen en diferentes etapas e tipos de respostas inmunitarias.
Control autónomo de refuxio de inmunidade
O papel dun circuíto reflexo do sistema nervioso autónomo colinérxico na regulación das respostas inmunes periféricas tamén aparece destacado 46. O vago é o principal nervio parasimpático que conecta o tronco cerebral cos órganos viscerais. O traballo de Kevin Tracey e outros apuntan a respostas antiinflamatorias xeneralizadas potentes en choque séptico e endotoxemia, desencadeadas por unha actividade do nervio vagal eferente que leva á supresión dos macrófagos periféricos 47-49. O vago activa as neuronas do ganglio celíaco adrenérxico periférico que inervan o bazo, o que leva á liberación de acetilcolina augas abaixo, que se une aos receptores nicotínicos alfa-7 dos macrófagos do bazo e do tracto gastrointestinal. Isto induce a activación da vía de sinalización SOCS2 JAK3 / STAT3, que suprime poderosamente a transcrición TNF-alfa 47. O ganglio celíaco adrenérxico tamén se comunica directamente cun subconxunto de células T de memoria que producen acetilcolina, que suprimen os macrófagos inflamatorios 48.
As células T asesinas naturais invariantes (iNKT) son un subconxunto especializado de células T que recoñecen lípidos microbianos no contexto de CD1d en lugar de antíxenos peptídicos. As células NKT son unha importante poboación de linfocitos implicados no combate a patóxenos infecciosos ea regulación da inmunidade sistémica. As células NKT residen e circulan principalmente a través da vasculatura e sinusoides do bazo e do fígado. Os nervios beta-adrenérxicos simpáticos no fígado sinalan directamente para modular a actividade celular NKT 50. Durante un modelo de accidente cerebrovascular (MCAO) de rato, por exemplo, a movilidad da célula NKT do fígado foi visiblemente suprimida, que foi revertida por denervación simpática ou antagonistas beta-adrenérxicos. Ademais, esta actividade inmunosupresora das neuronas noradrenérxicas nas células NKT provocou un incremento da infección sistémica e da lesión pulmonar. Por conseguinte, os sinais eferentes das neuronas autonómicas poden mediar unha potente inmunoespresión.

Insight do Dr. Alex Jimenez
A inflamación neurogénica é unha resposta inflamatoria local xerada polo sistema nervioso. Crese que desempeña un papel fundamental na patoxénese dunha variedade de problemas de saúde, incluíndo, enxaqueca, psoriase, asma, fibromialxia, eczema, rosácea, distonía e sensibilidade química múltiple. Aínda que a investigación inflamatoria neurogénica asociada ao sistema nervioso periférico foi ampliamente investigada, o concepto de inflamación neurogénica dentro do sistema nervioso central aínda necesita máis investigacións. Segundo varios estudos de investigación, con todo, as deficiencias de magnesio son as principais causas da inflamación neurogénica. O seguinte artigo mostra unha visión xeral dos mecanismos de inflamación neurogénica no sistema nervioso, o que pode axudar aos profesionais da saúde a determinar a mellor forma de tratamento para coidar unha variedade de problemas de saúde relacionados co sistema nervioso.
Conclusións
Cales son os respectivos roles específicos dos sistemas nerviosos somatosensorial e autonómico na regulación da inflamación e do sistema inmunitario (Fig. 4)? A activación dos nociceptores leva a reflexos axónicos locais, que localmente recrutan e activan células inmunes e, polo tanto, son principalmente proinflamatorios e espacialmente confinados. Pola contra, a estimulación autonómica leva a unha inmunosupresión sistémica ao afectar as pozas de células inmunes no fígado e no bazo. Os mecanismos de sinalización aferentes na periferia que conducen ao desencadeamento do circuíto reflexo colinérxico vagal inmunosupresor son mal comprendidos. Non obstante, o 80-90% das fibras vagais son fibras sensoriais aferentes primarias e, polo tanto, os sinais das vísceras, moitas potencialmente impulsadas por células inmunes, poden levar á activación de interneuronas no tronco cerebral e a través delas a unha saída en fibras vagais eferentes 46.
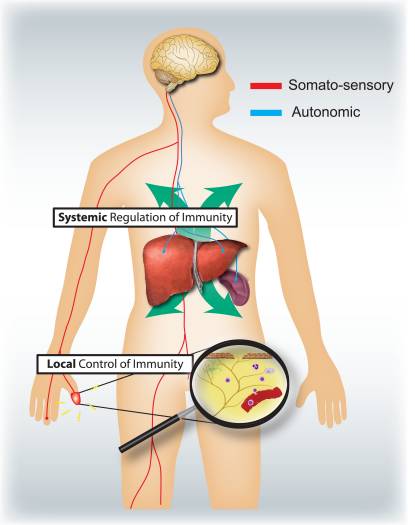
Imaxe 4: Os sistemas nerviosos sensoriais e autonómicos modulan respostas inmunitarias locais e sistémicas respectivamente. Os nociceptores que inervan superficies epiteliales (por exemplo, a pel e os pulmóns) induce respostas inflamatorias localizadas, activando mastocitos e células dendríticas. En inflamación de vías respiratorias alérxicas, dermatitis e artrite reumatoide, as neuronas nociceptoras desempeñan un papel na condución da inflamación. En contraste, os circuítos autonómicos que inervan os órganos viscerales (por exemplo o bazo eo fígado) regulan as respostas inmunitarias sistémicas mediante o bloqueo do macrófago ea activación da célula NKT. En accidente vascular cerebral e endotoxemia séptica, estas neuronas desempeñan un papel inmunosupresor.
Normalmente, o curso de tempo ea natureza da inflamación, xa sexa durante a infección, reaccións alérxicas ou patoloxías autoinmunes, están definidas polas categorías das células inmunitarias implicadas. Será importante saber que diferentes tipos de células inmunes están reguladas por sinais sensoriais e autonómicos. Unha avaliación sistemática do que os mediadores poden liberarse dos nociceptores e as neuronas autonómicas ea expresión dos receptores destes por diferentes células inmunitarias innatas e adaptativas poden axudar a resolver esta cuestión.
Durante a evolución, desenvolvéronse vías moleculares similares de detección de perigos tanto para a inmunidade innata como para a nocicepción, aínda que as células teñen liñaxes de desenvolvemento completamente diferentes. Mentres que os PRR e os canles iónicos nocivos con cancro de ligando son estudados por separado por inmunólogos e neurobiólogos, a liña entre estes dous campos é cada vez máis difusa. Durante o dano nos tecidos e a infección patóxena, é probable que a liberación de sinais de perigo conduza a unha activación coordinada tanto das neuronas periféricas como das células inmunes con comunicación bidireccional complexa e unha defensa integrada do hóspede. O posicionamento anatómico dos nociceptores na interface co ambiente, a velocidade da transducción neuronal e a súa capacidade para liberar cócteles potentes de mediadores de acción inmune permítelle ao sistema nervioso periférico modular activamente a resposta inmune innata e coordinar a inmunidade adaptativa augas abaixo. Pola contra, os nociceptores son moi sensibles aos mediadores inmunes, que activan e sensibilizan as neuronas. A inflamación neuroxénica e inmune mediada non son, polo tanto, entidades independentes senón que actúan xuntas como dispositivos de alerta precoz. Non obstante, o sistema nervioso periférico tamén xoga un papel importante na fisiopatoloxía, e quizais na etioloxía, de moitas enfermidades inmunes como asma, psoríase ou colite porque a súa capacidade para activar o sistema inmune pode amplificar a inflamación patolóxica 15-17. O tratamento para os trastornos inmunes pode ter que incluír, polo tanto, a orientación de nociceptores e tamén de células inmunes.
Grazas
Agradecemos ao NIH o soporte (2R37NS039518).
En conclusión,É esencial comprender o papel da inflamación neuroxénica á hora de defender a hospedaxe e a inmunopatoloxía para determinar o enfoque de tratamento axeitado para unha variedade de problemas de saúde do sistema nervioso. Ao ver as interaccións das neuronas periféricas coas células inmunes, os profesionais sanitarios poden avanzar en enfoques terapéuticos para axudar a aumentar a defensa do hóspede e suprimir a inmunopatoloxía. O obxectivo do artigo anterior é axudar aos pacientes a comprender a neurofisioloxía clínica da neuropatía, entre outros problemas de saúde de lesións nerviosas. Información referenciada polo Centro Nacional de Información sobre Biotecnoloxía (NCBI). O alcance da nosa información limítase á quiropráctica, así como ás lesións e condicións da columna vertebral. Para falar do asunto, non dubide en preguntarlle ao doutor Jiménez ou contactar connosco en 915-850-0900 .
Comisariado polo Dr. Alex Jiménez

Temas adicionais: dor nas costas
Dor nas costas é unha das causas máis frecuentes de discapacidade e días perdidos no traballo en todo o mundo. En realidade, a dor lumbar atribúese como o segundo motivo máis frecuente para as consultas médicas, superadas en número por infeccións respiratorias superiores. Aproximadamente o 80 por cento da poboación experimentará algún tipo de dor nas costas polo menos unha vez ao longo da súa vida. A columna vertebral é unha estrutura complexa composta de ósos, articulacións, ligamentos e músculos, entre outros tecidos brandos. Por iso, feridas e / ou condicións agravadas, como discos herniados, pode levar a síntomas de dor nas costas. As lesións deportivas ou as lesións por accidentes automovilísticos adoitan ser a causa máis frecuente de dor nas costas, con todo, ás veces o movemento máis sinxelo pode ter resultados dolorosos. Afortunadamente, as opcións de tratamento alternativas, como o coidado quiropráctico, poden axudar a aliviar a dor nas costas mediante o uso de axustes espiñentos e manipulacións manuais, mellorando o alivio da dor.

TEMA IMPORTANTE EXTRA: Manexo de baixa dor nas costas
MÁIS TEMAS: EXTRA EXTRA: Dor e tratamentos crónicos
En branco
References
Acordo pechado
Ámbito de práctica profesional *
A información aquí contenida en "O papel da inflamación neurogénica" non pretende substituír unha relación individual cun profesional da saúde cualificado ou un médico licenciado e non é un consello médico. Animámoslle a que tome decisións sobre a saúde baseándose na súa investigación e colaboración cun profesional sanitario cualificado.
Información do blog e debates de alcance
O noso ámbito de información limítase a quiropráctica, músculo-esqueléticos, medicamentos físicos, benestar, contribuíndo etiolóxico trastornos viscerosomáticos dentro de presentacións clínicas, dinámica clínica do reflexo somatovisceral asociado, complexos de subluxación, problemas de saúde sensibles e/ou artigos, temas e discusións de medicina funcional.
Proporcionamos e presentamos colaboración clínica con especialistas de diversas disciplinas. Cada especialista réxese polo seu ámbito profesional e a súa xurisdición de licenza. Usamos protocolos funcionais de saúde e benestar para tratar e apoiar a atención das lesións ou trastornos do sistema músculo-esquelético.
Os nosos vídeos, publicacións, temas, temas e coñecementos abarcan asuntos clínicos, cuestións e temas relacionados co noso ámbito de práctica clínica e apoian directa ou indirectamente o noso ámbito de práctica.*
A nosa oficina intentou razoablemente proporcionar citas de apoio e identificou o estudo ou estudos de investigación relevantes que apoian as nosas publicacións. Proporcionamos copias dos estudos de investigación de apoio dispoñibles para os consellos reguladores e o público logo de solicitude.
Entendemos que cubrimos asuntos que requiren unha explicación adicional de como pode axudar nun determinado plan de atención ou protocolo de tratamento; polo tanto, para debater máis sobre o tema anterior, non dubide en preguntar Dr. Alex Jiménez, DC, ou póñase en contacto connosco 915-850-0900.
Estamos aquí para axudarche a ti e á túa familia.
Bendicións
Dr. Alex Jiménez ANUNCIO, MSACP, RN*, CCST, IFMCP*, CIFM*, ATN*
e-mail: coach@elpasofunctionalmedicine.com
Licenciado como Doutor en Quiropráctica (DC) en Texas & Novo México*
Número de licenza de Texas DC TX5807, New Mexico DC Número de licenza NM-DC2182
Licenciada como enfermeira rexistrada (RN*) in Florida
Licenza Florida Licenza RN # RN9617241 (Nº de control 3558029)
Estado compacto: Licenza multiestatal: Autorizado para Practicar en Estados 40*
Dr. Alex Jimenez DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST
A miña tarxeta de visita dixital





