Equipo de Quiropráctica e Medicina Funcional da Clínica de Voltas. O noso corpo está nunha batalla constante e interminable pola supervivencia. As células nacen, as células destrúense. Os científicos estiman que cada célula debe soportar máis de 10,000 ataques individuais de especies reactivas do osíxeno (ROS) ou radicais libres. Sen falla, o corpo ten un incrible sistema de autocuración que resiste o ataque e reconstruíu o que foi danado ou destruído. Esta é a beleza do noso deseño.
Comprender a bioloxía do envellecemento e traducir os coñecementos científicos en intervencións que melloren a saúde da vida adulta mediante tratamentos. É útil ter unha visión clara e consensuada sobre o que constitúe exactamente un tratamento anti-envellecemento.
Desde antes dos días da busca de lonxevidade de Ponce de León, o home sempre foi atraído pola oportunidade da eterna xuventude. O coidado quiropráctico co seu movemento de saúde é un método poderoso para estabilizar e mellorar esta capacidade de autocuración. O doutor Alex Jiménez analiza os conceptos que rodean a pandora anti-envellecemento.
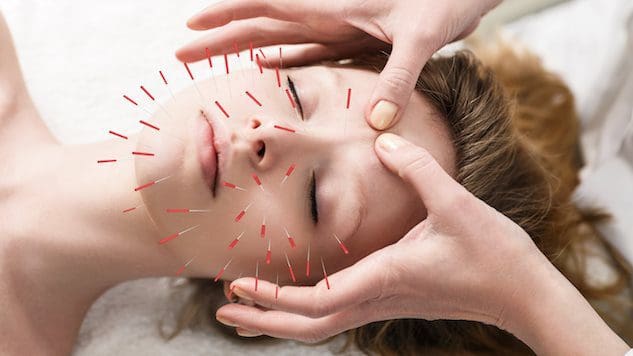
Para as persoas que queren mellorar ou manter a saúde da pel, pode incorporar a acupuntura axudar a mellorar a pel e combater o proceso de envellecemento?
Acupuntura cosmética
A acupuntura cosmética segue a práctica de acupuntura tradicional de inserción de agullas. O obxectivo é revertir os signos do envellecemento e mellorar a saúde da pel. Ás veces denomínase rexuvenecemento facial de acupuntura, que se utilizou como alternativa aos liftings cirúrxicos e outros procedementos convencionais. Os estudos preliminares examinaron como pode axudar a eliminar as manchas de idade, levantar as pálpebras caídas e reducir as engurras. (Younghee Yun et al., 2013)
Como funciona a acupuntura
Na medicina tradicional chinesa ou TCM, a acupuntura úsase durante moito tempo para mellorar o fluxo de enerxía - qi ou chi - por todo o corpo. Crese que esta enerxía circula por vías enerxéticas coñecidas como meridianos. Cando se producen problemas de saúde, segundo TCM, hai obstrucións ou bloqueos na circulación.
Os acupuntores poden restaurar a circulación/fluxo óptimo e mellorar a saúde introducindo agullas en puntos de acupuntura específicos. (Institutos Nacionais de Saúde, 2007)
Acupuntura cosmética
Dise que a acupuntura cosmética mellora a saúde da pel e actúa como un tratamento anti-envellecemento estimulando a produción de coláxeno. Esta proteína é un compoñente principal da pel. A capa interna da pel perde coláxeno e firmeza a medida que o corpo envellece. Non obstante, necesítanse máis investigacións para apoiar a afirmación de que a acupuntura pode promover a produción de coláxeno. Algúns suxiren que a acupuntura cosmética axuda a rexuvenecer a pel mellorando a enerxía global do corpo. Un estudo descubriu que os individuos viron melloras despois de cinco sesións de acupuntura cosmética facial. (Younghee Yun et al., 2013) Non obstante, recoméndase realizar dez tratamentos unha ou dúas veces por semana para obter resultados óptimos. Despois diso, os tratamentos de mantemento realízanse cada catro a oito semanas. A diferenza do Botox ou dos recheos dérmicos, a acupuntura cosmética non é unha solución rápida. O foco é crear cambios a longo prazo na pel e no corpo, o que significa mellorar:
Cando as agullas se introducen na pel, crean feridas coñecidas como microtraumatismos positivos. As capacidades naturais de curación e reparación do corpo actívanse cando detecta estas feridas. Estas puncións estimulan os sistemas linfático e circulatorio, que entregan nutrientes e osíxeno ás células da pel, nutíndoas de dentro para fóra.
Isto axuda a igualar a tez e favorece o brillo da pel.
Os microtraumas positivos tamén estimulan a produción de coláxeno.
Isto axuda a mellorar a elasticidade, minimizando liñas e engurras.
Alternativas
Varios remedios naturais poden axudar a mellorar a saúde da pel e ofrecer beneficios contra o envellecemento. As ceramidas son unha molécula de graxa que se atopa naturalmente na capa superior da pel e un ingrediente que se usa nos produtos para o coidado da pel. Estes poden protexer contra a sequedade da pel relacionada co envellecemento. (L Di Marzio 2008) A investigación preliminar suxire que a aplicación de té branco na pel pode loitar contra a descomposición do coláxeno e a elastina, unha proteína que apoia a elasticidade da pel e evita a flacidez. Tamén hai probas de que substancias naturais como o aceite de argán, o aceite de borraxa e o espinheiro poden ofrecer beneficios hidratantes que poderían mellorar a pel.Tamsyn SA Thring et al., 2009)
Aínda que se necesitan máis probas de acupuntura cosmética, a integración da acupuntura pode axudar a xestionar o estrés e mellorar a saúde xeral. As persoas que estean considerando a acupuntura cosmética deben consultar co seu médico de atención primaria para ver se é adecuada para eles.
Mellorando a saúde xuntos: abrazando a avaliación e o tratamento multidisciplinares
References
Yun, Y., Kim, S., Kim, M., Kim, K., Park, JS e Choi, I. (2013). Efecto da acupuntura cosmética facial na elasticidade facial: un estudo piloto de etiqueta aberta e dun só brazo. Medicina alternativa e complementaria baseada na evidencia: eCAM, 2013, 424313. doi.org/10.1155/2013/424313
O Centro Nacional de Medicina Complementaria e Alternativa. (2007). Acupuntura: unha introdución. Páxina web do Centro Nacional de Medicina Complementaria e Alternativa. choimd.com/downloads/NIH-info-on-acupuncture.pdf
Kuge, H., Mori, H., Tanaka, TH e Tsuji, R. (2021). Fiabilidade e validez da folla de verificación facial (FCS): lista de verificación para a autosatisfacción coa acupuntura cosmética. Medicamentos (Basilea, Suíza), 8(4), 18. doi.org/10.3390/medicines8040018
Di Marzio, L., Cinque, B., Cupelli, F., De Simone, C., Cifone, MG e Giuliani, M. (2008). Aumento dos niveis de ceramida cutánea en suxeitos anciáns tras unha aplicación tópica a curto prazo de esfingomielinase bacteriana de Streptococcus thermophilus. Revista internacional de inmunopatoloxía e farmacoloxía, 21 (1), 137–143. doi.org/10.1177/039463200802100115
Thring, TS, Hili, P. e Naughton, DP (2009). Actividades anticolaxenasas, antielastasas e antioxidantes de extractos de 21 plantas. Medicina alternativa e complementaria BMC, 9, 27. doi.org/10.1186/1472-6882-9-27
Manter a columna vertebral dun individuo en plena forma equivale a menos dor e máis mobilidade, flexibilidade e liberdade. O corpo desgástase e é un efecto natural do envellecemento que sucede con cada un de nós. Os problemas da columna vertebral relacionados co envellecemento poden ser graves se non se abordan e non se adoptan con exercicios, estiramentos e mantemento quiropráctico.
Envellecemento e costas
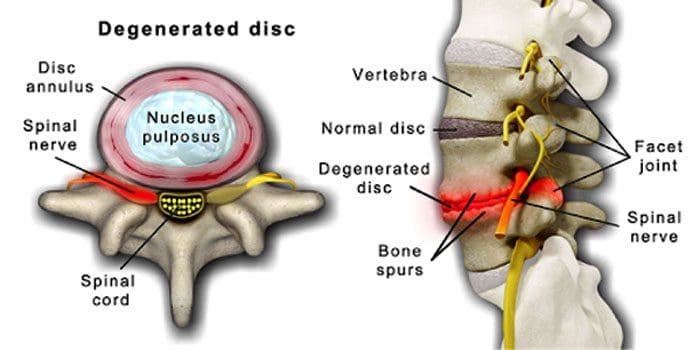
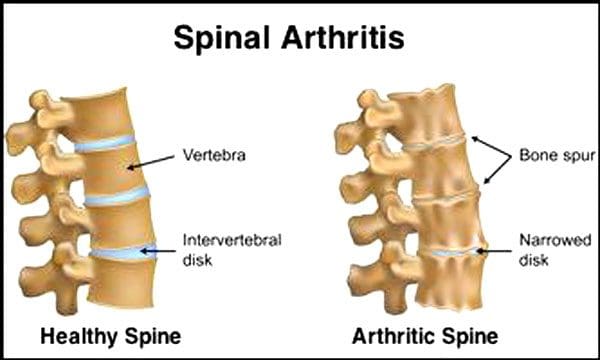
É normal que os discos e articulacións espiñais se deterioren coa idade. A estenose espinal ou o estreitamento da canle espinal tamén poden formar parte do proceso de envellecemento. Hai dúas condicións provocadas polo envellecemento enfermidade de disco degenerativa artrite que tamén pode incluír endurecemento dos ligamentos da columna vertebral e osteoporose.
A enfermidade dexenerativa do disco experimenta o 40% dos individuos de 40 anos
Aumenta ata o 80% para individuos de 80 anos ou máis.
Centrase arredor discos que gradualmente pasan de ser principalmente auga a principalmente graxa.
Cando está gordo, os discos redúcense e perden elasticidade.
Os Centros de Control e Prevención de Enfermidades din isto O 23% dos adultos estadounidenses teñen artrite. Esta é unha afección que afecta principalmente ás articulacións das facetas. As articulacións están inchadas, o que reduce o rango de movemento e pode afectar aos nervios espiñais, causando dor, debilidade e ciática. Co tempo os ligamentos ao redor e na columna vertebral endurécense, reducindo o rango de movemento, provocando estenose. A perda ósea ou osteoporose prodúcese por cambios nas hormonas e outros factores como a nutrición. O envellecemento é un proceso natural, pero os individuos poden axudar ás súas espiñas a manterse en plena forma non importa a idade que teñan.
Practicar unha postura sa
De inmediato adecuada mecánica corporal sa é imprescindible. Manterse consciente e consciente da postura corporal mantén o aliñamento e mantén o corpo equilibrado. Unha postura sa contribuirá a reducir os efectos de:
Estenose espinal
Enfermidade de disco degenerativa
Herniación
Risco de fracturas da columna vertebral
Practicar unha postura adecuada inclúe:
Reducir o slouching
Asegúrese de que a estación de traballo estea en plena forma e sexa ergonomicamente sólida
Asegúrese de dobrar os xeonllos ao levantar e manteña a columna vertebral o máis vertical posible.
Ioga
Ioga pode ser altamente beneficioso para unha columna vertebral máis sa e xuvenil. O ioga cumpre tres áreas para manter a columna vertebral en plena forma. Isto inclúe:
Exercicio regular
Mantén a flexibilidade
Alcanza o peso corporal ideal
O ioga é unha actividade que desafía a idade para a columna vertebral. Porque:
Mantén a forza
Flexibilidade
Postura
Saldo
Pode ser útil para unha variedade de enfermidades da columna vertebral, especialmente a dor de artrite
As caídas poden causar feridas graves. O ioga tamén pode axudar a traballar no equilibrio.
Vexa un quiropráctico
A medicina preventiva é fundamental para manter o corpo saudable, xuvenil e o máis forte posible. Un exame quiropráctico pode determinar se hai problemas na columna vertebral e un diagnóstico para desenvolver un plan de tratamento óptimo. Se a función corporal é limitada debido á dor nas costas e / ou nas pernas, póñase en contacto coa clínica de quiropraxia médica de lesións médicas e recuperación da columna vertebral en plena forma.
Composición do corpo
Rizos de bola de exercicio / estabilidade
Este exercicio traballa grupos musculares específicos da forza da columna vertebral e inclúe:
isquiotibiais
Glúteos
Abdominais profundos
Secuestradores e rotadores de cadeira
Exercicios coma este son un dos xeitos máis eficaces de construír forza e resistencia nos isquiotibiais, nas cadeiras e previr lesións. Para facer este adestramento:
Acuéstese de costas cos xeonllos dobrados
Levante as pernas para que a parte inferior dos pés se apoie sobre unha bola de exercicio
Estire as pernas ata que estean rectas
Manteña a posición por un segundo ou dous
Volve á parte superior do movemento mentres apertas os isquiotibiais
Traballar estes músculos axudará a que os movementos en cuclillas, lanzamentos ou flexións sexan máis doados na columna vertebral.
Descargo de responsabilidade sobre a publicación do blog do doutor Alex Jiménez
O alcance da nosa información limítase a medicamentos quiroprácticos, musculoesqueléticos, físicos, benestar e problemas de saúde sensibles e / ou artigos, temas e discusións de medicina funcional. Usamos protocolos de saúde e benestar funcionais para tratar e apoiar a atención a lesións ou trastornos do sistema músculo-esquelético. As nosas publicacións, temas, temas e ideas abarcan cuestións clínicas, problemas e temas que relacionan e apoian directa ou indirectamente o noso ámbito clínico de práctica. *
A nosa oficina fixo un intento razoable de proporcionar citas de apoio e identificou o estudo ou estudos de investigación pertinentes que apoian as nosas publicacións. Tamén poñemos copias dos estudos de investigación de apoio dispoñibles para o consello e / ou o público logo de solicitude. Entendemos que cubrimos asuntos que requiren unha explicación adicional sobre como pode axudar nun determinado plan de atención ou protocolo de tratamento; polo tanto, para discutir máis sobre o asunto anterior, non dubide en preguntar ao doutor Alex Jiménez ou contactar connosco no 915-850-0900. Proveedor (s) con licenza en Texas e Novo México *
Os alimentos que comemos poden ter un beneficio ou prexudicial para a nosa saúde. Unha mala alimentación pode causar unha variedade de problemas de saúde, incluída a obesidade, enfermidades cardiovasculares e diabetes tipo 2. Mentres tanto, unha nutrición adecuada pode facelo sentir energizado, reducir o risco de problemas de saúde, así como axudar a manter e regular un peso saudable. Se queres promover a lonxevidade, tes que alimentar o teu corpo con bos alimentos. No artigo seguinte, enumeraremos varios bos alimentos que ao final poden axudar a promover a lonxevidade, axudando tamén a mellorar a saúde e o benestar en xeral.
Vexetais Cruciferos
Os vexetais crucíferos teñen a capacidade única de cambiar as nosas hormonas, desencadear o sistema natural de desintoxicación do corpo e incluso reducir o crecemento das células cancerosas. Estes deben mastigarse ben ou comerse rallados, picados, zumados ou mesturados para liberar as súas propiedades beneficiosas. Tamén se descubriu que o sulforapano, que se atopa en vexetais crucíferos, axuda a protexer a parede dos vasos sanguíneos da inflamación que pode causar enfermidades cardíacas. Os vexetais crucíferos, como a col, o repolo, a col de Bruxelas, a coliflor e o brócoli son varios dos alimentos máis ricos en nutrientes do mundo.
Ensaladas Verdes
As verduras de folla cru teñen menos de 100 calorías por libra, o que os converte no alimento perfecto para a perda de peso. Comer máis verduras de ensalada tamén se asociou co risco reducido de ataque cardíaco, vertedura, diabetes e varios tipos de cancro. As verduras de folla crúa tamén son ricas no folato esencial de vitamina B, ademais da luteína e a zeaxantina, carotenoides que poden axudar a protexer os ollos. Os fitoquímicos solubles en graxa, como os carotenoides, que se atopan en verdes de ensalada como leituga, espinaca, kale, verdura de collard e verdes de mostaza tamén teñen efectos antioxidantes e antiinflamatorios no corpo.
Noces
As noces son un alimento con baixo contido glicémico e unha gran fonte de graxas saudables, proteínas vexetais, fibras, antioxidantes, fitosteroles e minerais, o que tamén axuda a reducir a carga glicémica de toda unha comida, converténdose nunha parte esencial dun anti-diabete. dieta. Independentemente da súa densidade calórica, comer froitos secos pode axudar a promover a perda de peso. As porcas tamén poden reducir o colesterol e axudar a reducir o risco de enfermidades cardíacas.
Sementes
As sementes, como as noces, tamén fornecen graxas saudables, antioxidantes e minerais. Non obstante, estas teñen máis proteínas e son ricas en minerais. As sementes de liño, liño e cánabo son ricas en graxas omega-3. As sementes de liño, liño e sésamo son tamén ricos lignáns ou fitoestrógenos que loitan contra o cancro de mama. Ademais, as sementes de sésamo son ricas en calcio e vitamina E, e as sementes de cabaza son ricas en cinc.
Bagas
As bagas son froitas ricas en antioxidantes que poden axudar a promover a saúde cardíaca. Estudos de investigación onde os participantes comeron amorodos ou arándanos diariamente durante varias semanas reportaron melloras na presión arterial, colesterol total e LDL e incluso signos de estrés oxidativo. As bayas tamén teñen propiedades contra o cancro e demostrouse que axudan a previr a caída cognitiva asociada ao envellecemento.
Roma
O fitoquímico máis coñecido nas granadas, o punicalaxina, é o responsable de máis da metade da actividade antioxidante da froita. Os fitoquímicos de Roma teñen beneficios contra o cancro, cardioprotectores e saudables para o cerebro. Nun estudo de investigación, os adultos maiores que tomaron zume de Roma diariamente durante 28 días tiveron un mellor rendemento nunha proba de memoria en comparación cos que beberon unha bebida placebo.
Feixóns
Comer faba e outros legumes pode axudar a equilibrar o azucre no sangue, reducir o apetito e protexerse contra o cancro de colon. As fabas son un alimento contra a diabetes que pode axudar a promover a perda de peso porque son dixeridas lentamente, o que ralentiza o aumento de azucre no sangue despois dunha comida e axuda a previr as ansias de alimentos promovendo a saciedade. Descubriuse o risco de cancro de colon por comer faba e outras leguminosas dúas veces por semana. Comer faba e outros legumes, como feixón vermello, feixón negro, garavanzos, lentellas e chícharos divididos, tamén ofrece unha protección importante contra outros cancro.
Cogomelos
Comer cogomelos regularmente está asociado a un risco reducido de cancro de mama. Os cogomelos brancos e Portobello son especialmente beneficiosos contra o cancro de mama porque teñen inhibidores da aromatase ou compostos que inhiben a produción de estróxenos. Os cogomelos demostraron ter efectos antiinflamatorios e proporcionar unha actividade inmune das células inmunitarias, prevención de danos no ADN, ralentización do crecemento das células cancerosas e inhibición da anxioxénese. Os cogomelos deben cociñarse sempre que os cogomelos crus teñen un produto químicamente potencialmente canceríxeno coñecido como agaritina que se reduce significativamente ao cociñar.
Cebolas e allo
A cebola e o allo proporcionan beneficios cardiovasculares e do sistema inmunitario, así como efectos anti-diabéticos e anti-cancro. Tamén se asociaron a un menor risco de cancro gástrico e de próstata. A cebola e o allo son coñecidos polos seus compostos organosulfuros que axudan a previr o desenvolvemento de cancro mediante a desintoxicación dos axentes canceríxenos, diminuíndo o crecemento das células cancerosas e bloqueando a anxioxénese. A cebola e o allo tamén teñen altas concentracións de antioxidantes flavonoides que promoven a saúde, que teñen efectos antiinflamatorios que poden axudar a previr o cancro.
tomates
Os tomates son ricos nunha variedade de nutrientes, como licopeno, vitamina C e E, betacaroteno e antioxidantes flavonol. O licopeno pode axudar a protexer contra o cancro de próstata, os danos causados polos raios UV e? enfermidade cardiovascular. O licopeno absórbese mellor cando se cociñan os tomates. Unha cunca de salsa de tomate ten aproximadamente 10 veces a cantidade de licopeno que unha cunca de tomate cru e picado. Ten en conta tamén que os carotenoides, como o licopeno, son mellor absorbidos cando se acompañan de graxas saudables, así que goza dos teus tomates nunha ensalada con noces ou cun aderezo a base de noces para obter beneficios nutricionais extra.
Os alimentos que inxerimos poden ser beneficiosos ou prexudiciais para a nosa saúde. A mala nutrición pode causar diversos problemas de saúde, incluíndo obesidade, enfermidades cardiovasculares e diabetes tipo 2. Mentres tanto, unha nutrición adecuada pode facerche sentir enerxizado, reducir o risco de problemas de saúde e axudar a manter e regular un peso saudable. Se queres promover a lonxevidade, tes que alimentar o teu corpo con bos alimentos. Os bos alimentos tamén poden axudar a reducir a inflamación asociada a unha serie de problemas de saúde, incluíndo dor nas articulacións e artrite. Os profesionais sanitarios, como os quiroprácticos, poden ofrecer consellos sobre dieta e estilo de vida para axudar a promover a saúde e o benestar. No seguinte artigo, listaremos varios bos alimentos que finalmente poden axudar a promover a lonxevidade. - Dr. Alex Jiménez DC, CCST Insight
Zesty Zumo de remolacha
Servings: 1 Tempo de cocción: minutos 5-10
1 pomelo, pelado e cortado en rodajas
1 mazá, lavada e cortada en rodajas
1 remolacha enteira e follas se as ten, lavadas e cortadas en rodajas
Pomo de xenxibre de 1 polgada, aclarado, pelado e picado
Tome todos os ingredientes nun exprimidor de alta calidade. Mellor servido de inmediato.
Só unha cenoria proporciónalle toda a inxestión diaria de vitamina A
Si, comer só unha cenoria cocida de 80 g (2 onzas) dáche betacaroteno suficiente para que o teu corpo produza 1,480 microgramos (mcg) de vitamina A (necesaria para a renovación celular da pel). Iso é máis que a inxestión diaria recomendada de vitamina A nos Estados Unidos, que é duns 900 mcg. É mellor comer cenorias cocidas, xa que isto suaviza as paredes celulares permitindo que se absorba máis betacaroteno. Engadir alimentos máis saudables á túa dieta é unha boa forma de mellorar a túa saúde xeral.
O alcance da nosa información limítase a medicamentos quiroprácticos, musculoesqueléticos, físicos, benestar e problemas de saúde sensibles e / ou artigos, temas e discusións de medicina funcional. Usamos protocolos de saúde e benestar funcionais para tratar e apoiar a atención de lesións ou trastornos do sistema músculo-esquelético. As nosas publicacións, temas, temas e ideas abarcan asuntos clínicos, cuestións e temas que relacionan e apoian directa ou indirectamente o noso ámbito clínico de práctica. * A nosa oficina intentou razoablemente proporcionar citas de apoio e identificou o estudo de investigación ou estudos que apoian as nosas publicacións. Tamén poñemos copias dos estudos de investigación de apoio dispoñibles para o consello e / ou o público logo de solicitude. Entendemos que cubrimos asuntos que requiren unha explicación adicional sobre como pode axudar nun determinado plan de atención ou protocolo de tratamento; polo tanto, para discutir máis sobre o asunto anterior, non dubide en preguntar ao doutor Alex Jiménez ou contactar connosco en 915-850-0900. Proveedor (s) con licenza en Texas * e Novo México *
Comisariado polo doutor Alex Jimenez DC, CCST
Referencias:
Joel Fuhrman, MD. 10 mellores alimentos que podes comer para vivir máis tempo e estar saudable Goodwell Health, 6 de xuño de 2020, www.verywellhealth.com/best-foods-for-longevity-4005852.
Dowden, Angela. "O café é unha froita e outros feitos alimentarios incriblemente verdadeiros" Estilo de vida de MSN, 4 de xuño de 2020, www.msn.com/en-us/foodanddrink/did-you-know/coffee-is-a-fruit-and-other-unbelievably-true-food-facts/ss-BB152Q5q?li=BBnb7Kz&ocid =mailsignout#image=24.
Se estás experimentando algunha destas situacións, os seus péptidos de coláxeno poden ser baixos.
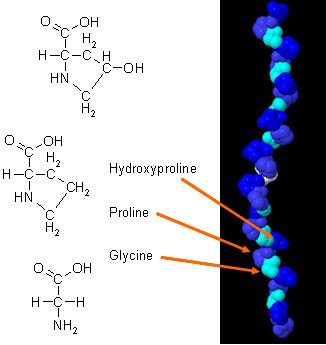
Hai estudos novos sobre como o coláxeno pode mellorar a composición corporal cando se combina con exercicios diarios. O coláxeno no corpo ten unha composición única de aminoácidos que desempeña un papel esencial na anatomía do corpo. A proteína de coláxeno é unha fonte concentrada de glicina, prolina e hidroxiprolina, e cando se compara con todas as outras proteínas da dieta, fai que o coláxeno sexa unha elección práctica potencial como proteína estrutural.
In un estudo 2015, os investigadores demostraron como os suplementos de coláxeno eficientes poden mellorar a composición corporal en homes activos. Os resultados mostran como cada individuo masculino está participando no adestramento do peso polo menos tres veces por semana e ten que complementarse con polo menos 15 gramos de péptidos de coláxeno para conseguir a máxima saúde. As avaliacións que proporciona a proba son proba de forza, análise de bioimpedancia (BIA) e biopsias musculares. Estas probas permiten que os individuos masculinos estean a bo rendemento despois de tomar os suplementos de coláxeno e os resultados mostran como a súa masa corporal tivo un aumento da masa corporal sen graxa. Outro estudo mostrou como a suplementación de proteína de coláxeno cando se combina con adestramento de resistencia que pode aumentar a masa muscular e a forza muscular tanto cos anciáns como coas persoas con sarcopenia.
Propiedades beneficiosas con coláxeno
Existen moitas propiedades beneficiosas que os suplementos de coláxeno poden proporcionar ao corpo cando se consume. Hai coláxeno hidrolizado e xelatina e poden axudar a mellorar a estrutura da pel dunha persoa. Aínda que non hai moitos estudos sobre suplementos de coláxeno, hai excelentes promesas para as áreas do corpo. Eles son:
Masa muscular: Os suplementos de coláxeno, combinados co adestramento de forza, poden aumentar a masa muscular e a forza no corpo.
Artrite: Os suplementos de coláxeno poden axudar ás persoas con artrose. Os estudos mostran que cando as persoas con artrosis toman suplementos de coláxeno, descubriron un descenso masivo na dor que estaban a sufrir.
Elasticidade da pel: En un estudo 2014, afirmou que as mulleres que tomaron suplementos de coláxeno e mostrou melloras na elasticidade da pel. O coláxeno tamén se pode usar en tratamentos tópicos para axudar a mellorar o aspecto da pel dunha persoa minimizando as liñas finas e as engurras.
Non só os suplementos de coláxeno proporcionan propiedades beneficiosas para as áreas específicas do corpo, senón que hai os catro tipos principais de coláxeno e cales son os seus roles no corpo humano e as súas funcións:
Introduza 1: O coláxeno tipo 1 tivo en conta o 90% do coláxeno do corpo e formado por fibras densamente envasadas que proporcionan estruturas á pel, ósos, tecidos conectivos e dentes do corpo.
Introduza 2: O coláxeno tipo 2 está composto por fibras pouco embaladas que se atopan na cartilaxe elástica, o que axuda a amortiguar as articulacións do corpo.
Introduza 3: O coláxeno tipo 3 axuda a apoiar a estrutura dos músculos, órganos e arterias que aseguran que o corpo funciona correctamente.
Introduza 4O coláxeno tipo 4 atópase nas capas da pel de todos e axuda coa filtración do corpo.
Dado que estes catro tipos de coláxeno están no corpo, é fundamental saber que o coláxeno pode diminuír naturalmente co paso do tempo coa idade, xa que o corpo producirá unha menor calidade de coláxeno. Un dos signos visibles de diminución de coláxeno é cando a pel do corpo humana se fai menos firme e suave, así como debilita a cartilaxe debido ao envellecemento.
Factores que poden danar o coláxeno
Aínda que o coláxeno pode diminuír naturalmente coa idade, moitos factores poden destruír colágenos prexudiciais para a pel. Os factores nocivos poden incluír:
Azucre e carbohidratos: Azucres e carbohidratos refinados pode interferir coa capacidade do coláxeno para repararse na pel. Polo tanto, ao minimizar o consumo de azucre e carbohidratos no corpo, pode reducir os efectos da disfunción do tecido vascular, renal e cutáneo.
Exposición ao sol: A pesar de obter o sol suficiente pode axudar a unha persoa a gozar do día, con todo, estando exposto ao sol durante un período prolongado pode causar danos para a pel e destruír os péptidos de coláxeno. Os efectos da sobreexposición do sol poden provocar que a pel envelleza a foto e produza estrés oxidativo no corpo.
fumador: Cando unha persoa fuma, pode reduce a produción de coláxeno no corpo, facendo que o corpo teña engurras prematuras e se o corpo está ferido, o proceso de curación será máis lento e pode provocar enfermidades no corpo.
Enfermidades autoinmunes: Algunhas enfermidades autoinmunes tamén poden danar a produción de coláxeno como o lupus.
Conclusión
O coláxeno é vital para o corpo, xa que axuda a que a pel sexa suave e firme. Naturalmente, diminuirá a medida que unha persoa envelleza, polo que tomar suplementos de coláxeno pode asegurarse de que o corpo poida funcionar correctamente. Cando os factores nocivos afectan ao corpo, poden deter ou incluso danar a produción de coláxeno e acelerar o proceso de formación das engurras prematuras, facendo que unha persoa pareza máis vella do que. Algunhas produtos pode axudar á actividade celular do corpo proporcionando unha estabilidade, biodisponibilidade e confort dixestivos máis excelentes.
O alcance da nosa información está limitado a problemas de saúde quiroprácticos, musculoesqueléticos e nerviosos ou artigos de medicina funcional, temas e discusións. Utilizamos protocolos de saúde funcionais para tratar lesións ou trastornos do sistema músculo-esquelético. A nosa oficina fixo un intento razoable para proporcionar citas de apoio e identificou o estudo ou estudos relevantes de apoio aos nosos postos. Tamén poñemos copias dos estudos de investigación de apoio para o consello e para o público previa solicitude. Para falar máis sobre o tema anterior, non dubide en preguntar ao doutor Alex Jiménez ou en contacto connosco 915-850-0900.
Referencias:
Bosch, Ricardo, et al. �Mecanismos de fotoenvellecemento e fotocarcinoxénese cutánea, e estratexias fotoprotectoras con fitoquímicos.� Antioxidantes (Basilea, Suíza), MDPI, 26 de marzo de 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4665475/.
Danby, William F. �Nutrición e envellecemento da pel: azucre e glicación.� Clínicas en Dermatoloxía, Biblioteca Nacional de Medicina dos Estados Unidos, 2010, www.ncbi.nlm.nih.gov/pubmed/20620757.
Jennings, Kerri-Ann. � Coláxeno: que é e para que serve?� Healthline, 9 de setembro de 2016, www.healthline.com/nutrition/collagen.
Jurgelewicz, Michael. �Un novo estudo demostra os beneficios dos péptidos de coláxeno para mellorar a composición corporal combinado co exercicio.� Deseños para a saúde, 31 de maio de 2019, blog.designsforhealth.com/node/1031.
Knuutinen, A, et al. �Fumar afecta a síntese de coláxeno e a rotación da matriz extracelular na pel humana.� O British Journal of Dermatology, Biblioteca Nacional de Medicina dos EUA, 2002 abril, www.ncbi.nlm.nih.gov/pubmed/11966688.
Proksch, E, et al. �A suplementación oral de péptidos específicos de coláxeno ten efectos beneficiosos na fisioloxía da pel humana: un estudo a dobre cego e controlado por placebo.� Farmacoloxía e Fisioloxía da pel, Biblioteca Nacional de Medicina dos Estados Unidos, 2014, www.ncbi.nlm.nih.gov/pubmed/23949208.
Schauss, Alexander G, et al. �Efecto do novo extracto de cartílago esternal de polo hidrolizado de baixo peso, coláxeno BioCell, na mellora dos síntomas relacionados coa osteoartrite: un ensaio aleatorizado, dobre cego e controlado por placebo.� Revista de Química Agraria e Alimentaria, Biblioteca Nacional de Medicina dos EUA, 25 X. 2012, www.ncbi.nlm.nih.gov/pubmed/22486722.
Zdzieblik, Denise, et al. �A suplementación de péptidos de coláxeno en combinación co adestramento de resistencia mellora a composición corporal e aumenta a forza muscular en homes sarcopénicos anciáns: un ensaio controlado aleatorizado. The British Journal of Nutrition, Cambridge University Press, 28 de outubro de 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4594048/.
Ao informar aos individuos sobre como a Universidade Nacional de Ciencias da Saúde proporciona o coñecemento para as xeracións futuras, a Universidade ofrece unha ampla variedade de profesións médicas para medicina funcional.
Como se lle diagnosticaron enfermidade celíaca, síndrome do intestino irritable, diverticulose / diveivertite ou síndrome de Leaky Gut?
Erupción excesiva, burping, ou inchazo?
¿Distensión anormal despois de certos probióticos ou suplementos naturais?
¿Sospeita de malabsorción nutricional?
Os problemas dixestivos paralizan coa relaxación?
Se estás experimentando algunha destas situacións, podes ter problemas intestinais e pode ter que probar o Programa 4R.
As sensibilidades alimentarias, a artrite reumatoide e a ansiedade asociaronse a unha permeabilidade gastrointestinal deteriorada. Estas diversas condicións poden pasar por moitos factores que poden afectar o tracto dixestivo. Se non se trata, pode ser o resultado dunha disfunción da barreira de permeabilidade intestinal, causando inflamacións e condicións de saúde graves que o intestino pode desenvolver. O programa 4R úsase para restaurar un intestino saudable no corpo e implica catro pasos. Son: eliminar, substituír, reinocular e reparar.
Permeabilidade intestinal
A permeabilidade intestinal axuda a protexer o corpo e asegúrase de que as bacterias nocivas non entren no intestino. Protexe o corpo de factores ambientais potenciais que pode ser prexudicial e está entrando polo tracto dixestivo. Pode ser toxina, microorganismos patóxenos e outros antíxenos que poidan danar o tracto dixestivo causando problemas. O revestimento intestinal está formado por unha capa de células epiteliais que están separadas por unións axustadas. Nun intestino sa, a estreita unión regula a permeabilidade intestinal permitindo que as substancias entren e viaxen a través da barreira intestinal e evitan que se absorban factores nocivos.
Algúns factores ambientais poden danar a unión axustada, e o resultado é que pode aumentar a permeabilidade intestinal, o que provoca hiperpermeabilidade intestinal ou goteo no corpo. Os factores que contribúen poden aumentar a permeabilidade intestinal como unha cantidade excesiva de graxas saturadas e alcol, deficiencias de nutrientes, estrés crónico e enfermidades infecciosas.
Cun aumento da permeabilidade intestinal no intestino, pode permitir que os antíxenos atravesen a mucosa do intestino e entren no torrente sanguíneo provocando unha resposta inmune e inflamación no corpo. Hai certas condicións gastrointestinais que están asociadas á hiperpermeabilidade intestinal e se non se trata, pode desencadear certas condicións autoinmunes que poden causar dano ao corpo.
Programa 4Rs
O 4Rs é un programa que os profesionais sanitarios aconsellan aos seus pacientes a usar cando están a tratar problemas dixestivos perturbadores e axudan a apoiar a cicatrización intestinal.
Eliminar o problema
O primeiro paso do programa 4Rs é eliminar patóxenos nocivos e desencadeantes de inflamación asociados ao aumento da permeabilidade intestinal. As tensións como o estrés e o consumo crónico de alcol poden causar moito dano ao corpo dun individuo. Así que dirixir estes factores nocivos ao corpo é tratalo con medicamentos, antibióticos, suplementos e recoméndase a eliminación de alimentos inflamatorios da dieta, incluíndo:
- Alcohol
- Glute
- Aditivos alimentarios
- Amidóns
- Algúns ácidos graxos
- Determinados alimentos cos que unha persoa é sensible
Substitución dos nutrientes
O segundo paso do programa 4Rs é substituír os nutrientes que están causando os problemas do intestino pola inflamación. Algúns nutrientes poden axudar a reducir a inflamación no intestino e asegurarse de que o tracto dixestivo está sendo soportado. Hai algúns alimentos antiinflamatorios nutritivos. Estes inclúen:
- Alimentos ricos en fibra
- Omega-3s
- Aceite de oliva
- Cogumelos
- Herbas antiinflamatorias
Hai certos suplementos que se poden usar para apoiar a función dixestiva axudando e absorbendo os nutrientes para promover un intestino saudable. O que fan as enzimas dixestivas é que axudan a descompoñer graxas, proteínas e carbohidratos no intestino. Isto axudará a beneficiar a persoas que presenten un trastorno dixestivo deteriorado, intolerancias alimentarias ou que teñan enfermidade celíaca. Suplementos como os suplementos de ácido biliar poden axudar á absorción de nutrientes combinando lípidos. Estudos afirmaron que os ácidos biliares se empregaron para tratar o fígado, a vesícula biliar e o conduto biliar, previndo a formación de cálculos biliares despois da cirurxía bariátrica.
Reinformado The Gut
O terceiro paso é o programa 4rs para reinocular o microbo intestino con bacterias beneficiosas para promover unha función intestina sa. Mostráronse estudos que os suplementos probióticos usáronse para mellorar o intestino recuperando bacterias beneficiosas. Con estes suplementos, proporcionan o intestino unha mellora secretando substancias antiinflamatorias no corpo, axudan a apoiar o sistema inmunitario, alterando a composición microbiana do corpo e reducindo a permeabilidade intestinal no sistema intestino.
Desde se atopan probióticos nos alimentos fermentados e considéranse transitorios xa que non son persistentes no tracto gastrointestinal e son beneficiosos. Sorprendente, aínda teñen un impacto na saúde humana debido a influír no intestino producindo vitaminas e compostos anti-microbianos, proporcionando así a diversidade e a función intestinal.
Reparación do Gut
O último paso do programa 4Rs é arranxar o intestino. Este paso consiste en reparar o revestimento intestinal do intestino con nutrientes e herbas específicas. Estas herbas e suplementos poden axudar a diminuír a permeabilidade intestinal e a inflamación no corpo. Algunhas destas herbas e suplementos inclúen:
- Aloe vera
- Goma mástica de Chios
- DGL (alcaçuz degriçrinizada)
- Raíz de malvavisco
- L-glutamina
- Omega-3s
� Polifenois
- Vitamina D
- Zinc
Conclusión
Xa que moitos factores poden afectar negativamente o sistema dixestivo dun xeito nocivo e poden contribuír a varias condicións de saúde. O obxectivo principal do programa 4Rs é minimizar estes factores que danan o intestino e reducir a inflamación e aumentar a permeabilidade intestinal. Cando o paciente está introducido nos factores beneficiosos que proporcionan os 4R, pode levar a un intestino san e sanado. Algunhas produtos están aquí para axudar ao sistema gastrointestinal apoiando os intestinos, mellorando o metabolismo do azucre e dirixindo os aminoácidos que están destinados a apoiar os intestinos.
O alcance da nosa información está limitado a problemas de saúde quiroprácticos, musculoesqueléticos e nerviosos ou artigos de medicina funcional, temas e discusións. Utilizamos protocolos de saúde funcionais para tratar lesións ou trastornos do sistema músculo-esquelético. A nosa oficina fixo un intento razoable para proporcionar citas de apoio e identificou o estudo ou estudos relevantes de apoio aos nosos postos. Tamén poñemos copias dos estudos de investigación de apoio para o consello e para o público previa solicitude. Para falar máis sobre o tema anterior, non dubide en preguntar ao doutor Alex Jiménez ou en contacto connosco 915-850-0900.
Referencias:
De Santis, Stefania, et al. �Claves nutricionais para a modulación da barreira intestinal Fronteiras en Inmunoloxía, Frontiers Media SA, 7 X. 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4670985/.
Ianiro, Gianluca, et al. �Suplemento enzimático dixestivo en enfermidades gastrointestinais.� Metabolismo das drogas actual, Bentham Science Publishers, 2016, www.ncbi.nlm.nih.gov/pmc/articles/PMC4923703/.
Mu, Qinghui, et al. �O intestino permeable como sinal de perigo para as enfermidades autoinmunes.� Fronteiras, Frontiers, 5 May 2017, www.frontiersin.org/articles/10.3389/fimmu.2017.00598/full.
Rezac, Shannon, et al. �Os alimentos fermentados como fonte dietética de organismos vivos.� Fronteiras en Microbioloxía, Frontiers Media SA, 24 2018 agosto, www.ncbi.nlm.nih.gov/pmc/articles/PMC6117398/.
Sander, Guy R., et al. �A interrupción rápida da función da barreira intestinal pola gliadina implica unha expresión alterada das proteínas da unión apical.� FEBS Prensa, John Wiley & Sons, Ltd, 8 de agosto de 2005, febs.onlinelibrary.wiley.com/doi/full/10.1016/j.febslet.2005.07.066.
Sartor, R Balfour. �Manipulación terapéutica da microflora entérica nas enfermidades inflamatorias do intestino: antibióticos, probióticos e prebióticos.� Gastroenteroloxía, Biblioteca Nacional de Medicina dos EUA, maio 2004, www.ncbi.nlm.nih.gov/pubmed/15168372.
A dor crónica é un problema de saúde común que afecta a moitas persoas nos Estados Unidos. Aínda que varias afeccións médicas, como a fibromialxia e a síndrome da dor miofascial, poden causar dor crónica, tamén pode desenvolverse debido a outros problemas de saúde. Estudos de investigación descubriron que a inflamación xeneralizada é a principal causa de dor crónica. A inflamación é un mecanismo natural de defensa contra lesións, enfermidades ou infeccións. Pero, se o proceso inflamatorio continúa demasiado tempo, pode chegar a ser problemático.
A inflamación sinala ao sistema inmunitario para curar e reparar o tecido danado, así como para protexerse contra bacterias e virus. Con todo, como se mencionou anteriormente, a inflamación crónica pode causar unha serie de problemas de saúde, incluídos síntomas de dor crónica. As modificacións de estilo de vida saudables poden axudar a controlar a dor crónica, pero primeiro entendemos as causas comúns da dor crónica.
Que é a inflamación aguda?
A inflamación aguda, por exemplo, ocorre tras unha lesión ou algo tan sinxelo coma unha dor de garganta. É unha resposta natural con efectos adversos, o que significa que funciona localmente na rexión onde se atopa o problema de saúde. Os signos comúns de inflamación aguda inclúen inchazo, enrojecimiento, calor, dor e perda de funcións, como indica a Biblioteca Nacional de Medicina. Cando se desenvolve a inflamación aguda, os vasos sanguíneos se dilatan facendo que o fluxo de sangue aumente e os glóbulos brancos na rexión lesionada promoven a recuperación.
Durante a inflamación grave, os tecidos danados liberan compostos chamados citocinas. As citocinas actúan como "sinais de emerxencia" que provocan as propias células inmunes do corpo humano, así como hormonas e numerosos nutrientes para solucionar o problema de saúde. Ademais, as substancias similares ás hormonas, coñecidas como prostaglandinas, provocan que os coágulos de sangue curen os tecidos danados e tamén poden provocar febre e dor como parte do procedemento inflamatorio. A medida que se recuperan os danos ou lesións, a inflamación diminúe.
Que é a inflamación crónica?
A diferenza da inflamación aguda, a inflamación crónica ten efectos a longo prazo. A inflamación crónica, tamén coñecida como inflamación persistente, produce baixos niveis de inflamación en todo o corpo humano, como demostra un aumento dos marcadores do sistema inmune situados nos tecidos sanguíneo e celular. A inflamación crónica tamén pode causar a progresión de varias enfermidades e condicións. Ás veces os niveis elevados de inflamación poden desencadearse aínda que non haxa lesións, enfermidades ou infeccións, que tamén poden facer que o sistema inmune reaccione.
Como resultado, o sistema inmune do corpo humano podería comezar a atacar células, tecidos ou órganos sans. Os investigadores seguen intentando comprender as consecuencias da inflamación crónica no corpo humano e os mecanismos implicados neste proceso de defensa natural. A modo de exemplo, a inflamación crónica asociouse a unha serie de problemas de saúde, como enfermidades cardíacas e ictus.
Unha teoría suxire que cando a inflamación permanece nos vasos sanguíneos, pode incentivar a acumulación de placa. Segundo a American Heart Association ou a AHA, se o sistema inmune identifica a placa como un invasor estranxeiro, as células brancas do sangue poden tentar amortiguar a placa atopada no sangue que flúe polas arterias. Isto pode crear un coágulo de sangue que poida bloquear o fluxo sanguíneo cara ao corazón ou ao cerebro, facendo que se volva inestable e se rompa. O cancro é outro problema de saúde asociado coa inflamación crónica. Ademais, segundo o National Cancer Institute, o dano no ADN tamén pode ser causado pola inflamación crónica.
A inflamación persistente e de baixa calidade con frecuencia non presenta síntomas, pero os profesionais sanitarios poden comprobar se hai unha proteína C reactiva ou CRP, coñecida como ácido lipoico, un marcador da inflamación que se atopa no sangue. Os niveis elevados de PCR están asociados cun maior risco de enfermidades cardiovasculares. Pódense atopar niveis elevados de CRP en trastornos crónicos como o lupus ou a artrite reumatoide.
No caso doutras enfermidades crónicas, como a fibromialxia, o sistema nervioso reacciona de forma excesiva á estimulación específica, pero é a inflamación a que causa síntomas de dor crónica. Subxectivamente, é case imposible distinguir a dor crónica causada por un sistema nervioso demasiado sensible e a dor crónica causada por unha inflamación xeneralizada. Ademais de buscar pistas no torrente sanguíneo, a nutrición, os hábitos de vida e as exposicións ambientais dunha persoa tamén poden promover a inflamación crónica.
A inflamación é o mecanismo de defensa natural do sistema inmunitario contra lesións, enfermidades ou infeccións. Aínda que esta resposta inflamatoria pode axudar a curar e reparar tecidos, a inflamación crónica e xeneralizada pode causar unha serie de problemas de saúde, incluídos síntomas de dor crónica. Un equilibrio a nutrición, incluída unha variedade de dietas e o xaxún, pode axudar a reducir a inflamación. O xaxún, tamén coñecido como restrición calórica, promove a apoptose celular e a recuperación mitocondrial. A dieta que imita o xaxún, que forma parte do plan de dieta de lonxevidade, é un programa dietético que "engana" ao corpo humano a un estado de xaxún para experimentar os beneficios do xaxún tradicional. Antes de seguir calquera das dietas descritas neste artigo, asegúrese de consultar a un médico.
Dr. Alex Jimenez DC, CCST Insight
Nutrición, dietas, xaxún e dor crónica
As dietas antiinflamatorias consisten principalmente en comer froitas e verduras frescas, peixes e graxas. O plan de dieta mediterránea, por exemplo, é unha dieta antiinflamatoria que promove comer cantidades moderadas de noces, inxerindo moi pouca carne e bebendo viño. As partes antiinflamatorias dos alimentos, como os ácidos graxos omega-3, protexen o corpo humano contra a damago provocado pola inflamación.
Unha dieta antiinflamatoria implica tamén estar lonxe de alimentos que poidan promover a inflamación. É ideal para diminuír a cantidade de alimentos que comeza con graxas trans e saturadas, como carnes. Ademais, unha dieta antiinflamatoria limita o consumo de hidratos de carbono e alimentos refinados, como o pan eo arroz. Isto tamén promove o corte da utilización da margarina e os aceites que son embalados con ácidos graxos omega-6, como o xirasol, o cártamo aceites de millo.
O xaxún ou a restricción calórica é coñecida por moito tempo que diminúe o estrés oxidativo e diminúe os mecanismos de envellecemento en varios organismos. Os efectos do xaxún implican a morte celular programada ou a apoptose, a transcrición, a eficiencia enerxética móbil, a biogênese mitocondrial, os mecanismos antioxidantes e o ritmo circadiano. O xaxún tamén contribúe á autofaxia mitocondrial, coñecida como mitofágia, onde os xenes das mitocondrias estimúlanse para someterse a apoptose, que promove a recuperación mitocondrial.
O xaxún intermitente pode axudarche a combater a inflamación, mellorar a dixestión e aumentar a túa lonxevidade. O corpo humano está deseñado para poder sobrevivir durante longos períodos de tempo sen comida. Estudos de investigación demostraron que o xaxún intermitente pode ter cambios positivos na composición xeral da microbiota intestinal. Ademais, o xaxún intermitente pode reducir a resistencia á insulina e aumentar a resposta do sistema inmunitario. Finalmente, o xaxún intermitente pode promover a produción dunha substancia, coñecida como β-hidroxibutirato, que bloquea unha parte do sistema inmune implicada en enfermidades inflamatorias, así como reducir substancialmente a produción de marcadores inflamatorios, como as citocinas e a proteína C reactiva. , ou CRP, mencionado anteriormente.
O Plan de dieta para a lonxevidade, presentado no libro polo doutor Valter Longo, elimina o consumo de alimentos procesados que poden causar inflamación, favorecendo o benestar e a lonxevidade. Este programa dietético único, a diferenza da maioría das dietas tradicionais, non promove a perda de peso. Aínda que pode experimentar redución de peso, o énfasis deste programa dietético único está en comer máis saudable. Demostrouse que o Plan de dieta para a lonxevidade axuda a activar a renovación baseada en células nai, reduce a graxa abdominal e prevén a perda ósea e muscular relacionada coa idade, así como a resistencia a enfermidades cardiovasculares, enfermidade de Alzheimer, diabetes e cancro.
A dieta de mimetismo en xaxún, ou FMD, permítelle experimentar os beneficios do xaxún tradicional sen privar o seu corpo de alimentos. A principal diferenza da fiebre aftosa é que no canto de eliminar por completo todos os alimentos durante varios días ou incluso semanas, só restrinxe a inxestión de calorías durante cinco días a partir de mes. A FMD pódese practicar unha vez ao mes para axudar a promover a saúde e o benestar en xeral.
Mentres calquera pode seguir a fiebre aftosa pola súa conta, o ProLon A dieta que imita o xaxún ofrece un programa de comida 5-día que foi embalado e etiquetado individualmente para cada día, que serve aos alimentos que necesita para a febre aftosa en cantidades e combinacións precisas. O programa de comidas está composto de alimentos listos para comer ou fáciles de preparar, con base en bares, sopas, merendas, suplementos, un concentrado de bebidas e tés. Antes de iniciar o programa Dieta que imita o xaxún ProLon, programa de comidas de 5 díasou calquera das modificacións do estilo de vida descritas anteriormente, por favor asegúrese de falar cun profesional da saúde para descubrir que tratamento da dor crónica é ideal para vostede.
O alcance da nosa información está limitado a problemas de quiropraxia, problemas de saúde espinal e artigos de medicina funcional, temas e discusións. Para seguir discutindo o tema anterior, póñase en contacto co Dr. Alex Jimenez ou póñase en contacto connosco 915-850-0900 .
Comisariado polo Dr. Alex Jiménez
Discusión de tema adicional: Dor aguda nas costas
Dor nas costas é unha das causas máis prevalentes de discapacidade e días perdidos no traballo en todo o mundo. A dor nas costas atribúese ao segundo motivo máis común das visitas a consultorios médicos, superado en número por infeccións respiratorias superiores. Aproximadamente o 80 por cento da poboación experimentará dores nas costas polo menos unha vez ao longo da súa vida. A columna vertebral é unha estrutura complexa composta de ósos, articulacións, ligamentos e músculos, entre outros tecidos brandos. Lesións e / ou condicións agravadas, como discos herniados, pode levar a síntomas de dor nas costas. As lesións deportivas ou as lesións por accidentes automovilísticos adoitan ser a causa máis frecuente de dor nas costas, con todo, ás veces o movemento máis sinxelo pode ter resultados dolorosos. Afortunadamente, as opcións de tratamento alternativas, como o coidado quiropráctico, poden axudar a aliviar a dor nas costas mediante o uso de axustes espiñentos e manipulacións manuais, mellorando o alivio da dor.
XIMOGENES Existen fórmulas profesionais exclusivas a través de profesionais de coidados de saúde con licenza selectiva. A venda por internet e o desconto das fórmulas XYMOGEN están estrictamente prohibidas.
Orgullosos, Dr. Alexander Jimenez fai que as fórmulas XYMOGEN estean dispoñibles só para os pacientes baixo o noso coidado.
Por favor, chame á nosa oficina para que poidamos asignar unha consulta médica para o acceso inmediato.
Se es paciente de Clínica médica e quiropráctica de lesións, pode preguntar sobre XYMOGEN chamando 915-850-0900.
Para a súa comodidade e revisión do XYMOGEN produtos, por favor revise a seguinte ligazón. *XYMOGEN-Catálogo-descargar
* Todas as políticas anteriores de XYMOGEN permanecen estrictamente en vigor.
A adhesión a unha dieta específica para manter unha nutrición adecuada ás veces fai que a alimentación sexa estresante. As modificacións naturais do estilo de vida son a clave para cambiar os teus hábitos alimentarios e isto pode axudarche a vivir unha vida máis longa e saudable. O Plan de Dieta para a Lonxevidade, creado polo Dr. Valter Longo, é unha selección de normas prácticas sobre comidas que se concentran en cambiar os seus patróns de alimentación para lograr unha saúde e benestar en xeral.
As regras do plan de dieta de longevidade
Simplemente seguindo as suxestións nutricionais que aparecen a continuación, pode revisar o plan de dieta actual e comezar a comer máis saudable sen o estrés dunha dieta tradicional. O Plan de Dieta para a Lonxevidade elimina o consumo de alimentos procesados que poden causar unha serie de problemas de saúde e aumenta o consumo de nutrientes que favorecen a lonxevidade. Este programa dietético único comparte os resultados de aproximadamente uns anos de estudos de investigación 25 todo nunha solución sinxela que pode axudar ás persoas a experimentar o benestar xeral a través dunha nutrición adecuada.
Non obstante, a diferenza da maioría das dietas tradicionais, o plan de dieta para a lonxevidade non promove a perda de peso. Aínda que pode experimentar redución de peso, o énfasis deste programa dietético único está en comer máis saudable. Demostrouse que o Plan de dieta para a lonxevidade axuda a activar a renovación baseada en células nai, adelgazar e reducir a graxa abdominal, previr a perda ósea e muscular relacionada coa idade, aumentar a resistencia ao desenvolvemento de enfermidades cardiovasculares, enfermidade de Alzheimer, diabetes e cancro. como estender a lonxevidade. A continuación, resumiremos os 8 consellos nutricionais máis comúns do plan de dieta para a lonxevidade que, finalmente, poden axudarche a facer a túa vida máis longa e saudable.
O Plan de Dieta de Lonxevidade é un programa dietético único deseñado polo Dr. Valter Longo para promover a saúde xeral, o benestar e a lonxevidade. A través de sinxelas modificacións no estilo de vida, a xente pode cambiar os seus hábitos alimentarios e aproveitar os moitos beneficios para a saúde deste programa dietético. Seguindo unha dieta pescatarian e seguindo a Dieta ProLon que imita o xaxún, entre os outros consellos nutricionais descritos a continuación, as persoas poden vivir unha vida máis longa e máis saudable. As dietas tradicionais a miúdo poden ser difíciles e estresantes, pero o plan de dieta de lonxevidade é un programa dietético único e práctico que pode ser adecuado para moitas persoas.
Dr. Alex Jimenez DC, CCST Insight
8 Consellos nutricionais do plan de dieta de longevidade
Siga unha dieta Pescatarian
Como parte do plan de dieta de lonxevidade, siga unha dieta pescatarian, que é case un centavo por cento en base a plantas e peixes. Ademais, asegúrese de limitar o consumo de peixe a dúas ou tres porcións cada semana, evitando os peixes con maior contido de mercurio, como atún, peixe espada, xurelo e fletán. Se tes máis de 100 e comeza a experimentar unha redución da masa muscular, forza e graxa, engade máis peixe á túa dieta, xunto con outros alimentos a base de animais, incluíndo ovos e queixos específicos, como feta ou pecorino, e iogur de cabra. leite.
Non comas demasiada proteína
Segundo o Plan de Dieta de Lonxevidade, debemos comer 0.31 a 0.36 gramos de proteína por quilo de graxa corporal todos os días. Se pesas 130lbs, debes comer 40 con 47 gramos de proteína por día, ou un equivalente de filetes de salmón 1.5, cunca 1 de garavanzos ou vasos de lentilas 2 1 / 2, dos cales o 30 gramos debe consumirse nunha comida. Se pesas 200 a 220lbs, debes comer 60 con 70 gramos de proteína ao día, ou un equivalente de dous filetes de salmón, 3 1 / 2 vasos de lentellas ou 1 1 / 2 cuncas de bico. O consumo de proteínas debe incrementarse despois da idade 65. Para a maioría de nós, basta un aumento por cento de 10 a 20 ou 5 a 10 cada día. Finalmente, a dieta de lonxevidade está exenta de proteínas animais como carnes vermellas, carnes brancas e aves de curral, con excepción das proteínas animais dos peixes. Este programa dietético único é máis ben comparado con proteínas vexetais como as leguminosas e as noces para optimizar a saúde e o benestar.
Aumente as graxas e carbohidratos complexos
Como parte do plan de dieta de lonxevidade, debes comer cantidades máis elevadas de graxas poliinsaturadas, como as que se atopan no salmón, améndoas, noces e aceite de oliva, mentres que debes comer cantidades máis baixas de graxas saturadas, hidrogenadas e trans. Do mesmo xeito, como parte do Plan de Dieta de Lonxevidade, tamén debe comer carbohidratos complexos, como os que se atopan no pan de trigo integral, legumes e verduras. Asegúrese de limitar o consumo de pastas, arroz, pan, froitas e zumes de froitas, que se poden converter en azucres cando cheguen ao intestino.
Tome suplementos dietéticos
O corpo humano necesita proteínas, ácidos graxos esenciais como omega-3 e omega-6, vitaminas, minerais e ata azucres para funcionar correctamente. Sempre que a inxestión de determinados nutrientes é demasiado baixa, os métodos de reparación, substitución e defensa do corpo humano poden diminuír ou parar, permitindo que os fungos, as bacterias e os virus causen danos que poidan provocar unha serie de problemas de saúde. Tome suplementos dietéticos con vitaminas e minerais, especialmente para os omega-3, segundo o recomendado polo profesional da saúde.
Coma varios alimentos da súa Asilvicultura
Para ter todos os nutrientes necesarios, cómpre comer unha gran variedade de alimentos, pero o mellor é elixir os alimentos comúns na táboa dos seus pais, avós e avós. Por exemplo, en moitos países do norte de Europa onde o leite foi consumido en xeral, a intolerancia á lactosa é relativamente rara, mentres que a intolerancia á lactosa é bastante común nos países do sur de Europa e asiáticos, onde o leite non era historicamente parte da dieta convencional dos adultos. Se unha persoa de ascendencia xaponesa residente nos Estados Unidos decide de súpeto comezar a beber leite, que probablemente non se serviu na mesa de cea dos seus avós, probablemente empezarán a sentirse mal. Os problemas máis comúns nestes casos son a intolerancia ou a autoinmunidades, como a resposta a alimentos ricos en glute como o pan e a pasta que se observan en persoas con enfermidade celíaca. Aínda que se precisan novas probas, é posible que as intolerancias alimentarias poidan estar relacionadas con moitas enfermidades autoinmunes, incluíndo a diabetes, a colite ea enfermidade de Crohn.
Coma dúas comidas ao día e un merenda
Segundo o Plan de dieta para a lonxevidade, é ideal tomar o almorzo e unha comida importante máis unha merenda nutritiva baixa en calorías e baixa en azucre todos os días. Mentres que para algunhas persoas pódese recomendar comer tres comidas e unha merenda todos os días. Moitas pautas nutricionais recomendan que debamos comer de cinco a seis comidas todos os días. Cando se recomenda ás persoas que coman con frecuencia, a miúdo pode facerse difícil regular a inxestión de calorías. Nos últimos vinte anos, aproximadamente o 70 por cento da poboación dos Estados Unidos considérase con sobrepeso ou obesidade. É moito máis difícil comer de máis no plan de dieta para a lonxevidade se só come dúas comidas e media todos os días. Necesitarían porcións masivas de legumes, verduras e peixe para alcanzar a cantidade que levaría ao aumento de peso. O alto alimento das comidas, máis a cantidade de comida, envíanlle ao estómago e ao cerebro un sinal de que xa ten comida suficiente. Este sistema de comidas importante ás veces pode ter que dividirse en dúas comidas para evitar problemas de dixestión. Os adultos e as persoas maiores propensas á perda de peso deben comer tres comidas ao día. Para as persoas que intentan perder peso, así como para as persoas con sobrepeso ou obesidade, o mellor consello nutricional sería o almorzo diario; cear ou xantar, pero non os dous, e substituír a comida perdida cunha merenda que conteña menos de 100 calorías e non máis de 3 a 5 g de azucre. Que comida omita depende do seu estilo de vida, con todo, non se recomenda saltar o almorzo debido aos seus problemas de saúde adversos. O beneficio de saltarse o xantar é máis tempo libre e enerxía. Pero hai un inconveniente para comer unha cea grande, especialmente para as persoas que sofren refluxo ácido ou problemas para durmir. Non obstante, o inconveniente de saltarse a cea é que pode eliminar a comida social do seu día.
Coma dentro dunha xanela de horas 12 cada día
Outro hábito común de comer adoptado por moitos centenarios é comer a tempo restrinxido ou limitar todas as comidas e bocadillos nunha xanela de 12 cada día. A eficiencia deste método demostrouse nos estudos de investigación en humanos e en animais. Xeralmente, xantaríase en 8 e logo xantar por 8 pm. Unha xanela máis curta de dez horas ou menos pode ser aínda mellor para a perda de peso, pero é bastante máis difícil de manter e pode aumentar o risco de desenvolver efectos secundarios, como cálculos biliares e incluso aumentar potencialmente a posibilidade de desenvolver enfermidades cardiovasculares. Non debe comer tres a catro horas antes de durmir.
Siga a dieta de imitación de xaxún ProLon
Persoas saudables menores de 10 anos deben seguir a Dieta ProLon Fasting Mimicking, programa de comidas de 5 días polo menos dúas veces ao ano. A febre aftosa é un dos principios clave promovidos polo Plan de Dieta de Lonxevidade. A dieta que imita o xaxún ofrece os mesmos beneficios para a saúde do xaxún sen xaxún. Ao comer 800 a calorías 1,100 en cantidades precisas e combinacións de alimentos que foron embalados e etiquetados individualmente por día, pode "enganar" ao corpo humano nun estado de xaxún. A través de varios estudos de investigación, o doutor Valter Longo descubriu que ao privar o corpo de alimentos deste xeito, as nosas células comezan a descompoñerse e rexenerar os nosos tecidos internos, a través dun proceso coñecido como autofagia, matando e substituíndo ou rexenerando as células danadas. Ademais, o xaxún pode revertir varios problemas de saúde, destruír as células canceríxenas e reducir significativamente a posibilidade de desenvolver a enfermidade de Alzheimer.
Co Plan de dieta para a lonxevidade presentado no libro polo doutor Valter Longo, comerás mellor, sentirásche mellor e, aínda que non está deseñado como un plan de adelgazamento, incluso podes perder algunhas libras. Non terás que considerar regras alimentarias complexas e tomar decisións difíciles con este programa dietético único. Despois de comprender estas modificacións de estilo de vida, poderás mellorar a túa saúde e benestar en xeral súa lonxevidade. o O alcance da nosa información está limitado a temas de quiropraxia, saúde vertebral e medicina funcional. Para discutir aínda máis o asunto, póñase en contacto con nós polo Dr. Alex Jimenez 915-850-0900 .
Comisariado polo Dr. Alex Jiménez
Discusión de tema adicional: Dor aguda nas costas
Dor nas costas é unha das causas máis prevalentes de discapacidade e días perdidos no traballo en todo o mundo. A dor nas costas atribúese ao segundo motivo máis común das visitas a consultorios médicos, superado en número por infeccións respiratorias superiores. Aproximadamente o 80 por cento da poboación experimentará dores nas costas polo menos unha vez ao longo da súa vida. A columna vertebral é unha estrutura complexa composta de ósos, articulacións, ligamentos e músculos, entre outros tecidos brandos. Lesións e / ou condicións agravadas, como discos herniados, pode levar a síntomas de dor nas costas. As lesións deportivas ou as lesións por accidentes automovilísticos adoitan ser a causa máis frecuente de dor nas costas, con todo, ás veces o movemento máis sinxelo pode ter resultados dolorosos. Afortunadamente, as opcións de tratamento alternativas, como o coidado quiropráctico, poden axudar a aliviar a dor nas costas mediante o uso de axustes espiñentos e manipulacións manuais, mellorando o alivio da dor.
XIMOGENES Existen fórmulas profesionais exclusivas a través de profesionais de coidados de saúde con licenza selectiva. A venda por internet e o desconto das fórmulas XYMOGEN están estrictamente prohibidas.
Orgullosos, Dr. Alexander Jimenez fai que as fórmulas XYMOGEN estean dispoñibles só para os pacientes baixo o noso coidado.
Por favor, chame á nosa oficina para que poidamos asignar unha consulta médica para o acceso inmediato.
Se es paciente de Clínica médica e quiropráctica de lesións, pode preguntar sobre XYMOGEN chamando 915-850-0900.
Para a súa comodidade e revisión do XYMOGEN produtos, por favor revise a seguinte ligazón. *XYMOGEN-Catálogo-descargar
* Todas as políticas anteriores de XYMOGEN permanecen estrictamente en vigor.
A ferramenta Find A Practitioner de IFM é a rede de referencia máis grande en Medicina Funcional, creada para axudar aos pacientes a localizar a profesionais da Medicina Funcional en calquera parte do mundo. Os profesionais certificados IFM aparecen en primeiro lugar nos resultados da busca, dada a súa ampla educación en Medicina Funcional