Volver Apoio Clínico de Neurofisioloxía Clínica. El Paso, TX. Chiropractor, o Dr. Alexander Jimenez discute neurofisioloxía clínica. O Dr. Jimenez explorará a importancia clínica e as actividades funcionais das fibras nerviosas periféricas, a medula espinal, o tronco cerebral e o cerebro no contexto dos trastornos visceral e musculoesquelético. Os pacientes obterán unha comprensión avanzada da anatomía, xenética, bioquímica e fisioloxía da dor en relación a varios síndromes clínicos. Engadiranse bioquímica nutricional relacionada coa nocicepción e dor. E salientarase a implementación desta información nos programas de terapia.
O noso equipo está moi orgulloso de traer ás nosas familias e pacientes feridos só protocolos de tratamento comprobados. Ao ensinar o benestar holístico completo como estilo de vida, tamén cambiamos non só a vida dos nosos pacientes, senón tamén as súas familias. Facemos isto para que poidamos chegar a tantos habitantes de El Paso que nos necesiten, sen importar os problemas de accesibilidade. Para respostas a calquera dúbida que poida ter, chame ao Dr. Jiménez ao 915-850-0900.
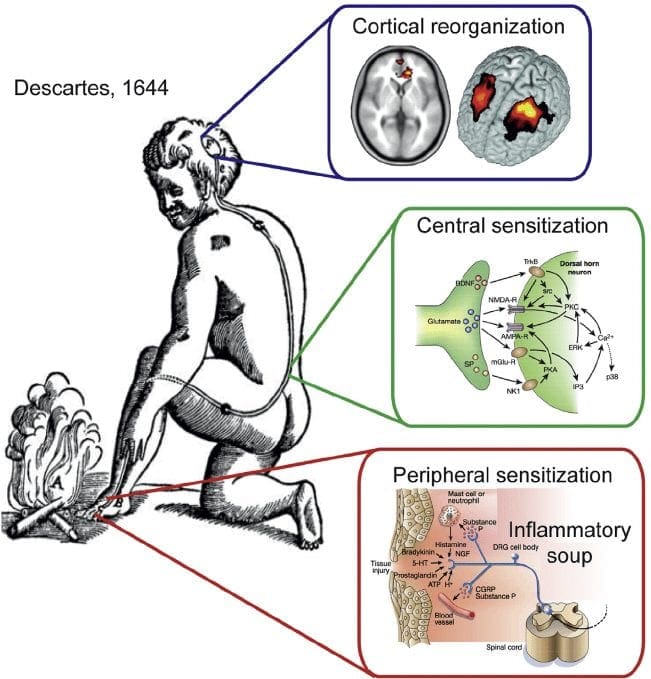
A dor neuropática é unha condición de dor crónica complexa que xeralmente está acompañada de lesións de tecidos brandos. A dor neuropática é común na práctica clínica e tamén supón un reto para pacientes e clínicos. Con dor neuropática, as propias fibras nerviosas poden estar danadas, non funcionais ou feridas. A dor neuropática é o resultado de danos causados por traumas ou enfermidades ao sistema nervioso periférico ou central, onde a lesión pode ocorrer en calquera sitio. Como resultado, estas fibras nerviosas danadas poden enviar sinais incorrectas a outros centros de dor. O efecto dunha lesión de fibras nerviosas consiste nun cambio na función neuronal, tanto na rexión da lesión coma no redor da lesión. Os signos clínicos de dor neuropática normalmente inclúen fenómenos sensoriais, como dor espontánea, parestesias e hiperalgesia.
A dor neuropática, definida pola Asociación Internacional do Estudo da Dor ou a IASP, é a dor iniciada ou causada por unha lesión primaria ou disfunción do sistema nervioso. Podería resultar de danos a calquera lado da neuraxia: sistema nervioso periférico, sistema nervioso espinal ou supraspinal. Os trazos que distinguen a dor neuropática doutros tipos de dor inclúen dor e signos sensoriais máis alá do período de recuperación. Caracterízase nos seres humanos por dor espontánea, alodinia ou a experiencia de estimulación non ferida como dolorosa e causalxia, ou dor de ardor persistente. A dor espontánea inclúe sensacións de "agujas e agullas", ardor, tiro, apuñalamento e dor paroxística ou dor de choque eléctrico, moitas veces asociada a disestèsias e parestesias. Estas sensacións non só alteran o aparello sensorial do paciente, senón tamén o benestar, estado de ánimo, atención e pensamento do paciente. A dor neuropática está formada por síntomas "negativos", como perda sensorial e sensacións de hormigueo e síntomas "positivos", como parestesias, dor espontánea e aumento da sensación de dor.
As condicións frecuentemente relacionadas coa dor neuropática poden clasificarse en dous grupos principais: dor debido a dano no sistema nervioso central e dor por danos ao sistema nervioso periférico. Os accidentes vasculares corticales e subcorticales, lesións traumáticas da medula espiñal, a xiringa-myelia e a syringobulbia, as neuralgias trigeminal e glossofaríngeas, as neoplásicas e outras lesións ocupantes do espazo son as condicións clínicas que pertencen ao antigo grupo. As neuropatías de compresión nerviosa ou de atrapamiento, neuropatías isquémicas, polineuropatias periféricas, plexopatías, compresión de raíz nerviosa, tocón post-amputación e dor nas extremidades faciais, neuralxia postherpética e neuropatías relacionadas co cancro son as condicións clínicas que pertencen ao último grupo.
Fisiopatoloxía da Dor Neuropática
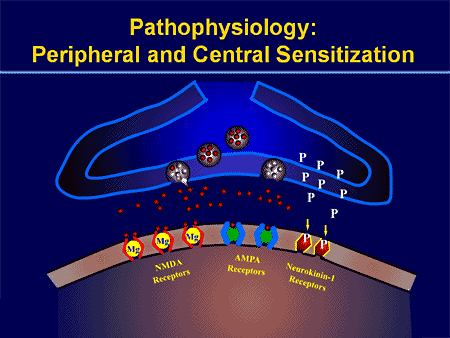
Os procesos e os conceptos fisiopatolóxicos subxacentes á dor neuropática son múltiples. Antes de cubrir estes procesos, unha revisión da circuitería de dor común é crítica. Os circuítos de dor regulares implican a activación dun nociceptor, tamén coñecido como o receptor da dor, en resposta a unha estimulación dolorosa. Unha onda de despolarización entrégase ás neuronas de primeira orde, xunto coa precipitación de sodio a través de canles de sodio e precipitación de potasio. As neuronas terminan no tronco cerebral no núcleo trigémino ou no corno dorsal da medula espiñal. É aquí onde o sinal abre canles de calcio con voltaxe na terminal pre-sináptica, permitindo que o calcio entre. O calcio permite que o glutamato, un neurotransmisor excitatorio, sexa liberado no área sináptica. O glutamato únese aos receptores de NMDA nas neuronas de segunda orde, provocando a despolarización.
Estas neuronas atravesan a medula espiñal e viaxan ata o tálamo, onde se sincronizan con neuronas de terceira orde. Estes se conectan ao sistema límbico e á corteza cerebral. Hai tamén un camiño inhibitorio que prevén a transmisión de sinal de dor desde o corno dorsal. As neuronas anti-nociceptivas orixinan a torsión cerebral e percorren a medula espiñal onde se sincronizan con interneuronas curtas no corno dorsal liberando dopamina e noradrenalina. As interneuronas modulan a sinapse entre a neurona de primeira orde e a neurona de segunda orde liberando o ácido aminotérmico gamma ou o GABA, un neurotransmisor inhibitorio. En consecuencia, o cese de dor é o resultado da inhibición das sinapses entre as neuronas de primeira e segunda orde, mentres que o aumento da dor pode ser o resultado da supresión das conexións sinápticas inhibitorias.
Non obstante, o mecanismo subxacente á dor neuropática non é tan claro. Varios estudos en animais revelaron que moitos mecanismos poden estar implicados. Non obstante, hai que recordar que o que se aplica ás criaturas non sempre se pode aplicar ás persoas. As neuronas de primeira orde poden aumentar o seu disparo se están parcialmente danadas e aumentan a cantidade de canles de sodio. As descargas ectópicas son consecuencia dunha despolarización mellorada en determinados sitios da fibra, o que causa dor espontánea e dor relacionada co movemento. Os circuítos inhibitorios poderían diminuír no nivel do corno dorsal ou nas células nai cerebral, así como os dous, permitindo que os impulsos da dor viaxen sen ser opostos.
Ademais, pode haber alteracións no procesamento central da dor cando, por mor da dor crónica e o uso de algunha droga e / ou medicamentos, as neuronas de segunda e terceira orde poden crear unha "memoria" de dor e sensibilizarse. Aumenta a sensibilidade das neuronas espinales e reduce os limiares de activación. Outra teoría demostra o concepto de dor neuropática simpaticamente mantida. Esta noción demostrouse por analxésia logo da simpatectomía de animais e persoas. Non obstante, unha mestura de mecánica pode estar implicada en moitas enfermidades neuropáticas ou mixtas crónicas somáticas e neuropáticas. Entre eses retos no campo da dor, e moito máis en canto a dor neuropática, é a capacidade de comprobar. Hai un compoñente dobre para isto: primeiro, avaliando a calidade, intensidade e avance; e segundo, diagnosticar correctamente a dor neuropática.
Non obstante, hai algunhas ferramentas de diagnóstico que poden axudar aos médicos a evaluar a dor neuropática. Para os principiantes, os estudos de condución nerviosa e os potenciales evocados por sensores poden identificar e cuantificar a extensión do dano ás vías sensoriais, pero non nociceptivas, mediante o seguimento das respostas neurofisiolóxicas aos estímulos eléctricos. Adicionalmente, a percepción dos pasos sensoriais de proba sensorial en reacción aos estímulos externos de intensidades variables aplicando a estimulación á pel. A sensibilidade mecánica aos estímulos táctiles vese meditada por ferramentas especializadas, como os xenes von Frey, pinprick con agullas entrelazadas, así como sensibilidade á vibración xunto con vibrameters e dor térmica con termodinámicos.
Tamén é moi importante realizar unha avaliación neurolóxica integral para identificar as disfuncións motoras, sensoriais e autonómicas. En definitiva, hai numerosos cuestionarios que se utilizan para distinguir a dor neuropática na dor nociceptiva. Algúns deles inclúen só consultas de entrevistas (por exemplo, o cuestionario neuropático e a dor de identificación), mentres que outros conteñen as preguntas da entrevista e as probas físicas (por exemplo, a escala de Leeds Evaluation of Neuropathic Symptoms and Signs) ea nova ferramenta exacta, a Avaliación Normalizada A dor, que combina seis preguntas de entrevistas e dez valoracións fisiolóxicas.
Modalidades de tratamento para a dor neuropática
Os regras farmacolóxicas apuntan aos mecanismos da dor neuropática. Non obstante, ambos tratamentos farmacolóxicos e non farmacolóxicos proporcionan alivio total ou parcial en case a metade dos pacientes. Moitos testemuños baseados na evidencia suxiren usar mesturas de medicamentos e / ou medicamentos para funcionar para o maior número posible de mecanismos. A maioría dos estudos investigaron principalmente neurálgicas post-herpéticas e neuropáticas diabéticas dolorosas, pero os resultados non se aplican a todas as condicións de dor neuropáticas.
Antidepresivos
Os antidepresivos aumentan os niveis de serotonina e norepinefrina sináptica, mellorando así o efecto do sistema analxésico descendente asociado á dor neuropática. Foron o alicerce da dor neuropática. As accións analxésicas poden ser atribuíbles ao bloqueo de adrenalina e de dopamina non-adrenalina, o que pode supoñer unha inhibición descendente, un antagonismo do receptor NMDA e un bloqueo de canles de sodio. Antidepresivos tricíclicos, como TCAs; por exemplo, amitriptilina, imipramina, nortriptilina e doxepina, son poderosos contra dor de dor continuo ou ardor xunto con dor espontánea.
Os antidepresivos tricíclicos demostraron ser significativamente máis efectivos para a dor neuropática que os inhibidores da recaptação de serotonina específicos, ou os ISRS, como a fluoxetina, paroxetina, sertralina e citalopram. A razón pode ser que inhiban a recaptación de serotonina e nor-epinefrina, mentres que os ISRs só inhiben a recaptación de serotonina. Os antidepresivos tricíclicos poden ter efectos secundarios desagradables, incluíndo náuseas, confusión, bloques de conducción cardíaca, taquicardia e arritmias ventriculares. Tamén poden causar aumento de peso, un reducido limite de convulsión e hipotensión ortostática. Os tricíclicos deben ser utilizados con coidado nos anciáns, que son particularmente vulnerables aos seus efectos secundarios agudos. A concentración do fármaco no sangue debe monitorearse para evitar a toxicidade en pacientes que son metabolizadores de medicamentos lentos.
Os inhibidores da recaptação da serotonina-noradrenalina ou SNRI son unha nova clase de antidepresivos. Do mesmo xeito que as TCAs, parecen ser máis efectivos que os ISRS para tratar a dor neuropática porque tamén inhiben a recaptação de nin-epinefrina e dopamina. A venlafaxina é tan efectiva contra as polineuropatías debilitadoras, como a neuropatía diabética dolorosa, como a imipramina, na mención de TCA e as dúas son significativamente superiores ao placebo. Do mesmo xeito que as TCA, as SNRI parecen conferir beneficios independientemente dos seus efectos antidepresivos. Os efectos secundarios inclúen sedación, confusión, hipertensión e síndrome de abstinencia.
Drogas antiepilépticas
As drogas antiepilépticas poden utilizarse como tratamento de primeira liña especialmente para certos tipos de dor neuropática. Actúan modulando as canles de calcio e de voltaxe por voltaxe, mellorando os efectos inhibitorios do GABA e inhibindo a transmisión excitatora glutaminérmica. Non se demostrou que os medicamentos antiepilépticos sexan eficaces para a dor aguda. Nos casos de dor crónica, as drogas antiepilépticas parecen ser efectivas só na neuralxia trigeminal. Carbamazepina é habitualmente empregada para esta condición. A gabapentina, que funciona inhibindo a función da canle de calcio a través de accións agonistas na subunidade delta de alfa-2 da canle de calcio, tamén é coñecida por ser eficaz para a dor neuropática. Non obstante, a gabapentina actúa de forma centralizada e pode causar fatiga, confusión e somnolencia.
Analgésicos non opiáceos
Hai unha falta de datos fortes que axudan a usar medicamentos antiinflamatorios non esteroides ou AINEs, en alivio da dor neuropática. Isto pode deberse á falta dun compoñente inflamatorio para aliviar a dor. Pero foron utilizados indistintamente con opioides como adjuvantes no tratamento da dor de cancro. Aínda se informou de complicacións, especialmente en pacientes con debilitacións graves.
Analgésicos opiáceos
Os analxésicos opioides son un tema de gran debate para aliviar a dor neuropática. Actúan inhibindo os impulsos centrais de dor ascendente. Tradicionalmente, a dor neuropática previamente foi observada como resistente aos opiáceos, na que os opiáceos son métodos máis adecuados para tipos de dor nociceptivos coronarios e somáticos. Moitos médicos impiden o uso de opioides para tratar a dor neuropática, en gran parte debido ás preocupacións sobre abuso de drogas, problemas de dependencia e regulación. Pero hai moitos ensaios que atoparon analxésicos opiáceos para ter éxito. Oxicodona foi superior ao placebo para aliviar a dor, a alodinia, mellorar o sono e a discapacidade. Os opiáceos de liberación controlada, segundo unha base programada, son recomendados para pacientes con dor constante para fomentar niveis constantes de analgesia, previr as flutuacións na glucosa no sangue e evitar eventos adversos asociados a unha dosificación máis alta. Máis comúnmente, as preparacións orais úsanse por mor da súa maior facilidade de uso e rendibilidade. As preparacións transdérmica, parenteral e rectal adoitan usarse en pacientes que non poden tolerar as drogas orais.
Anestésicos locais
Anestésicos de actuación próximos son atractivos porque, grazas á súa acción rexional, teñen efectos secundarios mínimos. Actúan pola estabilización das canles de sodio nos axóns das neuronas periféricas de primeira orde. Eles funcionan mellor se só hai lesións no nervio parcial e os canles de sodio en exceso recolectáronse. A lidocaína tópica é o representante mellor estudiado do curso para a dor neuropática. En concreto, o uso deste parche de lidocaína por 5 por neuralxia post-herpética causou a súa aprobación pola FDA. O parche parece funcionar mellor cando hai un funcionamento da nociceptor danado, pero mantido, do sistema nervioso periférico do dermatoma involucrado que demostra como alodinia. Debe configurarse directamente na área sintomática para 12 horas e eliminarse por outras horas 12 e pode usarse durante anos deste xeito. Ademais das reaccións locais da pel, adoitan ser ben toleradas por moitos pacientes con dor neuropática.
Drogas Varias
A clonidina, un agonista alfa-2, demostrou ser eficaz nun subconxunto de pacientes con neuropatía periférica diabética. Os cannabinoides descubriron que desempeñaron un papel na modulación experimental da dor en modelos animais e evidencia de que a eficacia acumúlase. Os agonistas selectivos de CB2 suprimen a hiperalalxia e a alodinia e normalizan os limiares nociceptivos sen inducir a analxésia.
Xestión da dor intervencionista
Os tratamentos invasivos poderían considerarse para pacientes con dor neuropática intratábel. Estes tratamentos inclúen inxeccións epidurales ou perineurales de anestésicos locais ou corticoides, implantación de métodos de entrega de fármacos epidurales e intratecal e inserción de estimuladores da medula espiñal. Estes enfoques están reservados para pacientes con dor neuropática crónica intratável que non controlaron a xestión médica conservadora e que tamén experimentaron unha avaliación psicolóxica completa. Nun estudo de Kim et al, demostrouse que un estimulador da medula espinal era eficaz no tratamento da dor neuropática da orixe das raíces nerviosas.
Insight do Dr. Alex Jimenez
Con dor neuropática, os síntomas da dor crónica prodúcense debido a que as fibras nerviosas están mal, disfuncionais ou feridas, generalmente acompañadas por danos e lesións no tecido. Como resultado, estas fibras nerviosas poden comezar a enviar sinais de dor incorrectos a outras áreas do corpo. Os efectos da dor neuropática causada por lesións nas fibras nerviosas inclúen modificacións na función nerviosa tanto no lugar da lesión como nas áreas ao redor da lesión. A comprensión da fisiopatoloxía da dor neuropática foi un obxectivo para moitos profesionais da saúde, para determinar de forma eficaz o mellor enfoque de tratamento para axudar a xestionar e mellorar os seus síntomas. Do uso de medicamentos e / ou medicamentos, para o coidado quiropráctico, o exercicio físico, a actividade física e a nutrición, pódense utilizar diversos enfoques de tratamento para axudar a aliviar a dor neuropática para as necesidades de cada individuo.
Intervencións adicionais para a dor neuropática
Moitos pacientes con dor neuropática procuran opcións de tratamento alternativas e complementarias para tratar a dor neuropática. Outros réximes coñecidos que se usan para tratar a dor neuropática son a acupuntura, a estimulación percutánea do nervio eléctrico, a estimulación do nervio eléctrico transcutánea, o tratamento cognitivo-comportamento, a imaxe automática graduada eo tratamento de apoio e o exercicio. Entre estes, con todo, o coidado quiropráctico é un tratamento de tratamento alternativo ben coñecido comúnmente usado para axudar a tratar a dor neuropática. Os coidados de quiropraxia, xunto coa modificación de fisioterapia, exercicio, nutrición e estilo de vida poden ofrecer alivio para síntomas de dor neuropática.
A quiropraxia
O que se sabe é que unha aplicación de xestión integral é crucial para combater os efectos da dor neuropática. Deste xeito, o coidado quiropráctico é un programa de tratamento integral que podería ser eficaz na prevención de problemas de saúde relacionados co dano nervioso. O coidado quiropraxia brinda asistencia a pacientes con diferentes condicións, incluíndo aqueles con dor neuropática. Os sufrimentos da dor neuropática a miúdo utilizan medicamentos non esteroideos antiinflamatorios ou AINEs, como ibuprofeno ou analgésicos de prescrición pesada para axudar a aliviar a dor neuropática. Estes poden proporcionar unha corrección temporal, pero necesitan un uso constante para controlar a dor. Isto invariabelmente contribúe a efectos secundarios nocivos e en situacións extremas, dependencia de medicamentos con receita.
A asistencia quiropráctica pode axudar a mellorar os síntomas da dor neuropática e mellorar a estabilidade sen estas desvantaxes. Unha visión como o coidado quiropráctico ofrece un programa individualizado deseñado para identificar a causa principal do problema. A través do uso de axustes espiñentos e manipulacións manuais, un quiropráctico pode corrixir coidadosamente calquera desalineación espiñal, ou subluxacións, que se atopan ao longo da columna vertebral, o que podería reducir as consecuencias do nervio provocando a realineación da espiña dorsal. Restaurar a integridade da columna é esencial para manter un sistema nervioso central de alto funcionamento.
Un Quiroprácticos tamén pode ser un tratamento a longo prazo para mellorar o seu benestar xeral. Ademais dos axustes espiñentos e as manipulacións manuais, un quiropráctico pode ofrecer asesoramento nutricional, como a prescripción dunha dieta rica en antioxidantes ou o deseño dunha terapia física ou programa de exercicios para combater as manifestacións de dor nas neuronas. Unha condición a longo prazo esixe un remedio a longo prazo, e nesta capacidade, un profesional sanitario especializado en lesións e / ou condicións que afectan o sistema músculo-esquelético e nervioso, como un médico de quiropraxia ou quiropraxia, pode ser inestimable ao traballar para medir o cambio favorable ao longo do tempo.
As técnicas de representación física, física e de movemento demostraron ser beneficiosas para o tratamento da dor neuropática. O coidado quiropraxia tamén ofrece outras modalidades de tratamento que poden ser útiles para a xestión ou mellora da dor neuropática. A terapia con láser de baixo nivel, ou LLLT, por exemplo, gañou un tremendo protagonismo como tratamento da dor neuropática. Segundo unha variedade de estudos de investigación, concluíuse que a LLLT tiña efectos positivos no control da analgesia para a dor neuropática. No entanto, requírense estudos de investigación adicional para definir protocolos de tratamento que resumen os efectos da terapia con láser de baixo nivel en tratamentos de dor neuropáticos.
O coidado quiropraxia tamén inclúe consellos nutricionais que poden axudar a controlar os síntomas asociados coa neuropatía diabética. Durante un estudo de investigación, demostrouse unha dieta baseada en baixo contido de graxa para mellorar o control glicémico en pacientes con diabetes tipo 2. Despois de preto de 20 semanas do estudo piloto, os individuos implicados informaron cambios no seu peso corporal e conductancia electroquímica da pel no pé foi informado de que mellorou coa intervención. O estudo de investigación suxeriu un valor potencial na intervención na dieta con baixo contido de graxa na dieta para a neuropatía diabética. Ademais, os estudos clínicos descubriron que a aplicación oral de L-threonato de magnesio é capaz de evitar así como restaurar os déficits de memoria asociados coa dor neuropática.
O coidado quiropraxia tamén pode ofrecer estratexias de tratamento adicionais para promover a rexeneración nerviosa. A modo de exemplo, recomendouse mellorar a regeneración dos axóns para axudar a mellorar a recuperación funcional despois da lesión do nervio periférico. A estimulación eléctrica, xunto co exercicio físico ou actividades físicas, atopou para promover a regeneración nerviosa despois dunha reparación nerviosa retardada en humanos e ratas, segundo recentes estudos de investigación. Tanto a estimulación eléctrica como o exercicio foron determinados en última instancia como prometedores de tratamentos experimentais para a lesión do nervio periférico que parecen estar listos para ser transferidos ao uso clínico. É posible que se precisen máis estudos de investigación para determinar completamente os efectos destes en pacientes con dor neuropática.
Conclusión
A dor neuropática é unha entidade multifacética sen pautas particulares para coidar. É mellor xestionalo usando un enfoque multidisciplinar. A xestión da dor require avaliación continua, educación do paciente, garantindo o seguimento e tranquilidade do paciente. A dor neuropática é unha enfermidade crónica que fai que a opción para o mellor tratamento sexa desafiante. O tratamento individualizador implica a consideración do impacto da dor sobre o benestar, a depresión e as discapacidades do individuo, xunto coa formación continua e a avaliación. Os estudos sobre a dor neuropática, tanto a nivel molecular como en modelos animais, son relativamente novos pero moi prometedores. Prevese moitas melloras nos campos básico e clínico da dor neuropática, polo que se abren as portas a novas ou melloradas modalidades de tratamento para esta enfermidade discapacitante. O alcance da nosa información limítase á quiropráctica, así como ás lesións e condicións da columna vertebral. Para falar do asunto, non dubide en preguntarlle ao doutor Jiménez ou contactar connosco en 915-850-0900 .
Comisariado polo Dr. Alex Jiménez
Temas adicionais: dor nas costas
Dor nas costas é unha das causas máis frecuentes de discapacidade e días perdidos no traballo en todo o mundo. En realidade, a dor lumbar atribúese como o segundo motivo máis frecuente para as consultas médicas, superadas en número por infeccións respiratorias superiores. Aproximadamente o 80 por cento da poboación experimentará algún tipo de dor nas costas polo menos unha vez ao longo da súa vida. A columna vertebral é unha estrutura complexa composta de ósos, articulacións, ligamentos e músculos, entre outros tecidos brandos. Por iso, feridas e / ou condicións agravadas, como discos herniados, pode levar a síntomas de dor nas costas. As lesións deportivas ou as lesións por accidentes automovilísticos adoitan ser a causa máis frecuente de dor nas costas, con todo, ás veces o movemento máis sinxelo pode ter resultados dolorosos. Afortunadamente, as opcións de tratamento alternativas, como o coidado quiropráctico, poden axudar a aliviar a dor nas costas mediante o uso de axustes espiñentos e manipulacións manuais, mellorando o alivio da dor.
Depresión de ansiedade da dorTodo o mundo experimentou dor, pero hai persoas con depresión, ansiedade ou ambas. Combina isto coa dor e pode ser bastante intenso e difícil de tratar. As persoas que sofren depresión, ansiedade ou ambas as dúas tenden a experimentar dor severa e a longo prazo máis que outras persoas.
O camiño ansiedadea depresión e a superposición da dor entre si vense en síndromes de dor crónicos e discapacitantes, isto é, dor lumbar, dores de cabeza, dores nerviosas e fibromialxia. Os trastornos psiquiátricos contribúen á intensidade da dor e tamén aumentan o risco de discapacidade.
Depresión:A (trastorno depresivo maior ou depresión clínica) é un trastorno do humor común pero grave. Provoca síntomas graves que afectan a sensación, pensamento e manexo das actividades diarias dun individuo, é dicir, durmir, comer e traballar. Para ser diagnosticado de depresión, os síntomas deben estar presentes durante polo menos dúas semanas.
Humor persistente triste, ansioso ou "baleiro".
Sentimentos de desesperanza, pesimistas.
Irritabilidade.
Sentimentos de culpa, inutilidade ou impotencia.
Perda de interese ou pracer en actividades.
Disminución da enerxía ou fatiga.
Mover ou falar lentamente.
Sentirse inquedo e ter problemas para estar parado.
Dificultade de concentrarse, recordar ou tomar decisións.
Dificultade para durmir, espertar e durmir demasiado.
Cambios de apetito e peso.
Pensamentos de morte ou suicidio e intentos de suicidio.
Dolores ou dores, dores de cabeza, cólicas ou problemas dixestivos sen unha causa física clara e / ou que non facilitan o tratamento.
Non todos os que están deprimidos experimentan todos os síntomas. Algúns experimentan só algúns síntomas, mentres que outros poden experimentar varios. Varios síntomas persistentes ademais do baixo estado de ánimo son necesarioPara un diagnóstico de depresión maior. A gravidade e frecuencia dos síntomas e a duración variarán dependendo do individuo e da súa enfermidade particular. Os síntomas tamén poden variar segundo o estadio da enfermidade.
DEPRESIÓN ANTIESTADORA DE DOLOR
Obxectivos:
Cal é a relación?
Cal é a neurofisioloxía detrás dela?
Cales son as consecuencias centrais?
Cambios cerebrais na dor
Figura 1 Vías cerebrais, rexións e redes implicadas na dor aguda e crónica
Davis, KD et al. (2017) Probas de imaxe cerebral para dor crónica: problemas e recomendacións médicas, legais e éticas Nat. Rev. Neurol. doi: 10.1038 / nrneurol.2017.122
DOLOR, ANXIDADE E DEPRESIÓN
Conclusión:
A dor, especialmente crónica, está asociada á depresión e ansiedade
Os mecanismos fisiolóxicos que levan á ansiedade e á depresión poden ser de natureza multifactorial
Cando o sistema sensorial está afectado por lesións ou enfermidades, os nervios dentro dese sistema non poden funcionar correctamente para transmitir sensacións e sentimentos ao cerebro. Isto contribúe a miúdo a unha sensación de entumecemento ou falta de sensación. Non obstante, en certos casos, cando este sistema está danado, as persoas poden experimentar dor na zona afectada.
A dor neuropática non comeza bruscamente nin se resolve rapidamente; é unha Dor crónica condición que provoca síntomas de dor persistentes. Para a maioría dos individuos, a intensidade dos seus síntomas pode diminuír ao longo do día. Aínda que se supón que a dor neuropática está relacionada con problemas de saúde dos nervios periféricos, como a neuropatía causada pola diabetes ou a estenose espinal, as feridas no cerebro ou na medula espiñal tamén poden causar dor neuropática crónica. A dor neuropática tamén se refire como dor nerviosa.
A dor neuropática pódese contrastar coa dor nociceptiva. A dor neuropática non se desenvolve en ningunha circunstancia específica nin estímulo externo, senón que os síntomas prodúcense simplemente porque o sistema nervioso pode non funcionar en consecuencia. De feito, os individuos tamén poden experimentar dor neuropática mesmo cando a parte do corpo ferida ou ferida non está realmente presente. Esta enfermidade chámase dor pantasma nos membros, que pode ocorrer en persoas despois de ter unha amputación.
A dor nociceptiva é xeralmente aguda e desenvólvese en resposta a unha circunstancia específica, como cando alguén experimenta unha lesión súbita, como martelar un dedo cun martelo ou estucar un pé cando anda descalzo. Ademais, a dor nociceptiva tende a desaparecer unha vez que o sitio afectado cura. O corpo contén células nerviosas especializadas, coñecidas como nociceptores, que detectan estímulos nocivos que poden danar o corpo, como calor ou frío extremo, presión, pellizco e exposición a produtos químicos. Estes sinais de advertencia son entón transmitidos ao longo do sistema nervioso cara ao cerebro, o que resulta nunha dor nociceptiva.
Cales son os factores de risco para a dor neuropática?
Todo o que contribúe a unha falta de función no sistema nervioso sensorial pode provocar dor neuropática. Polo tanto, os problemas de saúde causados pola síndrome do túnel do carpo ou condicións similares poden desencadear a dor neuropática. O trauma, que resulta en lesións nerviosas, pode causar dor neuropática. Outras condicións que poden predispoñer aos individuos a desenvolver a dor neuropática inclúen: diabetes, deficiencias de vitamina, cancro, VIH, vertedura, esclerose múltiple, tellas e mesmo algúns tratamentos contra o cancro.
Cales son as causas da dor neuropática?
Hai moitas causas polas que os individuos poden desenvolver dor neuropática. Pero a nivel celular, unha explicación é un aumento na liberación de certos receptores que indican que a dor, xunto cunha diminución da capacidade dos nervios para modular estes sinais, leva á sensación de dor orixinaria da rexión afectada. Ademais, na medula espiñal, a rexión que exerce signos dolorosos reorganízase cos correspondentes cambios nas hormonas e perda de corpos móbiles que funcionan normalmente. Estas alteracións resultan na percepción da dor en ausencia de estimulación externa. No cerebro, a capacidade de bloquear a dor pode verse afectada tras unha lesión, como un accidente cerebrovascular ou trauma dunha lesión. Ao pasar o tempo, ocorre un dano celular adicional e continúa a sensación de dor. A dor neuropática está relacionada tamén coa diabetes, o consumo de alcohol crónico, certos tipos de cancro, deficiencia de vitamina B, enfermidades, outras enfermidades relacionadas con nervios, toxinas e medicamentos específicos.
Cales son os síntomas da dor neuropática?
Contrariamente a outras condicións neurolóxicas, a identificación da dor neuropática pode ser un reto. Non obstante, poden estar presentes varias sinais obxectivas, se as hai. Os profesionais da saúde teñen que descifrar e traducir unha variedade de palabras que os pacientes usan para describir a súa dor. Os pacientes poden describir os seus síntomas como agudos, aburridos, quentes, fríos, sensibles, picantes, profundos, picantes, ardentes, entre unha variedade doutros termos descritivos. Ademais, algúns pacientes poden experimentar dor a través dun lixeiro toque ou presión.
Nun esforzo para axudar a identificar a cantidade de dor que poden sufrir os pacientes, a miúdo úsanse diferentes escalas. Pídese aos pacientes que valoren a súa dor de acordo cunha escala visual ou un gráfico numérico. Existen moitos exemplos de escalas de dor, como o que se mostra a continuación. Moitas veces, as imaxes de rostros que representan unha variedade de niveis de dor poden ser útiles cando os individuos toman un momento difícil describir a cantidade de dor que están experimentando.
Dor crónica e saúde mental
Para moitos, o impacto da dor crónica pode non estar limitado á dor; tamén pode influír negativamente no seu estado mental. Os novos estudos realizados por científicos da Universidade de Northwestern en Chicago poden explicar por que os individuos que padecen dor crónica tamén sofren problemas de saúde aparentemente independentes, como depresión, estrés, falta de sono e dificultade para concentrarse.
A avaliación demostrou que as persoas con dor crónica mostran diferentes rexións do cerebro sempre activas, máis específicamente, a área asociada ao humor e á atención. Esta acción continua rexeita as conexións nerviosas do cerebro e deixa aos pacientes con dor crónica en maior risco de problemas psicolóxicos. Os investigadores suxeriron que obter sinais de dor de forma constante podería provocar a reconfiguración mental que afecta negativamente á mente. O cableado obriga aos seus cerebros a dedicar recursos mentais de xeito diferente para facer fronte ás tarefas cotiás, desde as matemáticas, ata recordar unha lista de compras, sentirse feliz.
A conexión dor-cerebro rexistrouse ben, polo menos de xeito anecdótico, e moitos profesionais sanitarios din que viron de primeira man o xeito no que o estado mental do paciente pode baixar cando soporta dor crónica. Os equívocos sobre a conexión dor-cerebro poden xurdir da falta de evidencias de que a dor ten unha influencia medible e duradeira no cerebro. Os investigadores esperan que con investigacións adicionais sobre os mecanismos de como a dor crónica fai ás persoas máis susceptibles a trastornos do estado de ánimo, as persoas terán a capacidade de controlar mellor o seu benestar xeral.
Cultura e dor crónica
Moitas cousas contribúen á nosa forma de experimentar e expresar a dor, con todo, recentemente suxeriron recentemente investigadores que a cultura se relaciona directamente coa expresión da dor. A nosa educación e os valores sociais afectan a forma en que expresamos a dor e tamén a súa propia natureza, intensidade e lonxitude. Non obstante, estas variables non son tan obvias como os valores socio-psicolóxicos, como a idade e o sexo.
A investigación afirma que a dor crónica é un proceso polifacético e que a interacción simultánea entre fisiopatoloxía, factores cognitivos, afectivos, condutuais e socioculturais resúmese co que se coñece como experiencia de dor crónica. Xurdiu que a dor crónica experimentouse de xeito diferente entre pacientes de diversas culturas e etnias.
Algunhas culturas fomentan a expresión da dor, particularmente no sur do Mediterráneo e no Oriente Medio. Outros individuos suprímeno, como nas moitas leccións para os nosos fillos sobre como comportarse con bravura e non chorar. A dor recoñécese como parte da experiencia humana. Estamos aptos a asumir que a comunicación sobre a dor atravesará perfectamente as fronteiras culturais. Pero a xente con dor está suxeita ás formas que as súas civilizacións os adestraron para experimentar e expresar dor.
Ambos os individuos en dor e profesionais da saúde experimentan dificultades para comunicar dor nas fronteiras étnicas. Nunha cuestión como a dor, onde a comunicación eficaz pode ter implicacións de longo alcance para a atención médica, a calidade de vida e a supervivencia potencial, o papel da cultura na comunicación da dor segue sendo insuficientemente valorado. A dor persistente é un encontro multidimensional, composto formado por entrelazados e co-influencia de factores biolóxicos e psicosociais. Coñecer a culminación destes factores é fundamental para comprender as diferenzas na súa manifestación e xestión.
Como se diagnostica a dor neuropática?
O diagnóstico de dor neuropática depende dunha avaliación adicional da historia dun individuo. Se se sospeita que hai dano nervioso subxacente, entón pode xustificarse a análise dos nervios xunto coa proba. O medio máis común para avaliar se un nervio está lesionado ou non é o uso de medicamentos electrodiagnósticos. Esta subespecialidade médica utiliza técnicas de estudos de condución nerviosa con electromielografía (NCS / EMG). A avaliación clínica pode mostrar evidencias de perda de traballo e pode incluír a avaliación do tacto lixeiro, a capacidade de diferenciar a dor aguda e a capacidade de discernir a temperatura, así como a avaliación da vibración.
Despois de completar un completo exame clínico, podería planificarse a análise de electrodiagnóstico. Estes estudos son realizados por neurólogos e fisioterapeutas especialmente adestrados. Se se sospeita a neuropatía, debería realizarse unha caza de causas reversibles. Isto pode incluír a función do sangue para as deficiencias de vitaminas ou problemas da tiroide e estudos de imaxe para excluír unha lesión estrutural que afecta á medula espiñal. Dependendo dos resultados desta proba, pode haber un medio para diminuír a intensidade da neuropatía e posiblemente reducir a dor que sofre un paciente.
Desafortunadamente, en moitas condicións, incluso un bo control da causa subxacente da neuropatía non pode reverter a dor neuropática. Isto vese normalmente en pacientes con neuropatía diabética. En casos raros, pode haber signos de cambios no patrón de crecemento da pel e do cabelo nunha rexión afectada. Estas alteracións poden estar asociadas a cambios na transpiración. Se están presentes, estes cambios poden axudar a identificar a probable presenza de dor neuropática relacionada cunha enfermidade coñecida como síndrome da dor rexional complexa.
Insight do Dr. Alex Jimenez
A dor neuropática é unha enfermidade de dor crónica que xeralmente está asociada a danos directos ou lesións no sistema nervioso ou nos nervios. Este tipo de dor é diferente da dor nociceptiva ou da típica sensación de dor. A dor nociceptiva é unha sensación aguda ou súbita de dor que fai que o sistema nervioso envíe sinais de dor inmediatamente despois do trauma. Con dor neuropática, con todo, os pacientes poden experimentar dor de tiro e ardor sen danos directos nin lesións. Comprender as posibles causas da dor neuropática do paciente fronte a calquera outro tipo de dor pode axudar aos profesionais sanitarios a atopar mellores xeitos de tratar as condicións crónicas da dor.
Que é o tratamento para a dor neuropática?
Utilízanse varios medicamentos para tratar a dor neuropática. A maioría destes fármacos utilízanse fóra de etiqueta, o que significa que a FDA aprobou o medicamento para tratar diferentes condicións e entón recoñeceuse que era vantaxoso para tratar a dor neuropática. Os antidepresivos tricíclicos, como a amitriptilina, a nortriptilina e a desipramina, foron prescritos para a xestión da dor neuropática durante varios anos.
Algunhas persoas atopan que poden ser moi eficaces para aliviarlles. Outros tipos de antidepresivos teñen mostrado alivio. En determinados pacientes utilizáronse inhibidores selectivos da recaptación da serotonina ou SSRI, como paroxetina e citalopram, e outros antidepresivos, como venlafaxina e bupropión. Outro tratamento frecuente da dor neuropática incorpora medicamentos anti-agresión, incluíndo carbamazepina, fenitoína, gabapentina, lamotrigina e outros.
En casos agudos de neuropatía dolorosa que non responden aos intermediarios de primeira liña, as drogas que se usan normalmente para tratar as arritmias cardíacas poden ter algún beneficio; con todo, estes poden provocar efectos secundarios significativos e, a miúdo, hai que supervisalos de preto. Os medicamentos aplicados directamente sobre a pel poden ofrecer beneficios modestos a perceptibles para algúns pacientes. Entre as formas que se usan habitualmente inclúese lidocaína (en parche ou xel) ou capsaicina.
O tratamento da dor neuropática depende da causa subxacente. Se a causa é reversible, entón os nervios periféricos poden rexenerarse e a dor diminuirá; con todo, esta redución da dor pode levar varios meses a anos. Outras opcións de tratamento alternativas, incluíndo a atención quiropráctica e a terapia física, tamén se poden utilizar para axudar a aliviar a tensión e a tensión nos nervios, axudando finalmente a mellorar os síntomas dolorosos.
Que é o pronóstico para a dor neuropática?
Moitas persoas con dor neuropática poden obter algunha axuda, incluso cando a dor persiste. Aínda que a dor neuropática non é perigosa para un paciente, a presenza de dor crónica pode afectar negativamente á calidade de vida. Os pacientes con dor nerviosa crónica poden sufrir privacións do sono ou trastornos do estado de ánimo, incluíndo depresión, ansiedade e estrés, como se mencionou anteriormente. Debido á alopecia inherente e á falta de retroalimentación sensorial, os pacientes corren o risco de desenvolver lesións ou infeccións ou, sen sabelo, de provocar unha escalada da lesión presente. Polo tanto, é esencial buscar atención médica inmediata e seguir pautas específicas dirixidas por un profesional sanitario por seguridade e precaución.
Pode previr a dor neuropática?
A mellor forma de previr a dor neuropática é evitar o desenvolvemento ou a progresión da neuropatía. Supervisar e cambiar as opcións de estilo de vida, incluíndo a restrición do uso de alcol e tabaco; manter un peso saudable para diminuír a posibilidade de diabetes, enfermidades dexenerativas nas articulacións ou derrame cerebral; e ter unha gran forma ergonómica no traballo ou cando se practican afeccións para reducir o risco de lesións por estrés repetitivas son estratexias para diminuír a probabilidade de desenvolver neuropatía e potencial dor neuropática. Asegúrese de buscar atención médica inmediata no caso de que haxa síntomas asociados á dor neuropática para proceder ao enfoque de tratamento máis adecuado. Para falar do asunto, non dubide en preguntarlle ao doutor Jiménez ou contactar connosco en 915-850-0900 .
Comisariado polo Dr. Alex Jiménez
Temas adicionais: dor nas costas
Dor nas costas é unha das causas máis frecuentes de discapacidade e días perdidos no traballo en todo o mundo. En realidade, a dor lumbar atribúese como o segundo motivo máis frecuente para as consultas médicas, superadas en número por infeccións respiratorias superiores. Aproximadamente o 80 por cento da poboación experimentará algún tipo de dor nas costas polo menos unha vez ao longo da súa vida. A columna vertebral é unha estrutura complexa composta de ósos, articulacións, ligamentos e músculos, entre outros tecidos brandos. Por iso, feridas e / ou condicións agravadas, como discos herniados, pode levar a síntomas de dor nas costas. As lesións deportivas ou as lesións por accidentes automovilísticos adoitan ser a causa máis frecuente de dor nas costas, con todo, ás veces o movemento máis sinxelo pode ter resultados dolorosos. Afortunadamente, as opcións de tratamento alternativas, como o coidado quiropráctico, poden axudar a aliviar a dor nas costas mediante o uso de axustes espiñentos e manipulacións manuais, mellorando o alivio da dor.
Se o sistema sensorial se ve afectado por lesións ou enfermidades, os nervios dese sistema non poden funcionar na transmisión de sensacións ao cerebro. Isto pode provocar sensación de adormecemento ou falta de sensación. Nalgúns casos cando o sistema sensorial está lesionado, os individuos poden experimentar dor na rexión afectada. A dor neuropática non comeza rápido ou remata rapidamente. É unha enfermidade crónica que leva a síntomas de dor persistente. Para moitos, a intensidade dos síntomas pode vir e ir durante todo o día. A dor neuropática crese que está asociada a problemas nerviosos periféricos, é dicir, a neuropatía causada pola diabetes, a estenose da columna vertebral, a lesión no cerebro ou a medula espiñal tamén poden provocar dor neuropática crónica.
DOLOR NEUROPÁTICO
Obxectivos:
¿Que é iso?
Cal é a fisiopatoloxía detrás dela?
Cales son as causas?
Cales son algunhas das vías?
Como podemos solucionalo?
DOLOR NEUROPÁTICO
Dor iniciada ou causada por unha lesión primaria ou disfunción no sistema nervioso somatosensorial.
A dor neuropática adoita ser crónica, difícil de tratar e moitas veces resistente á xestión analxésica estándar.
PATOGENESIS DO DOLOR NEUROPÁTICO
MECANISMOS PERIFÉRICOS
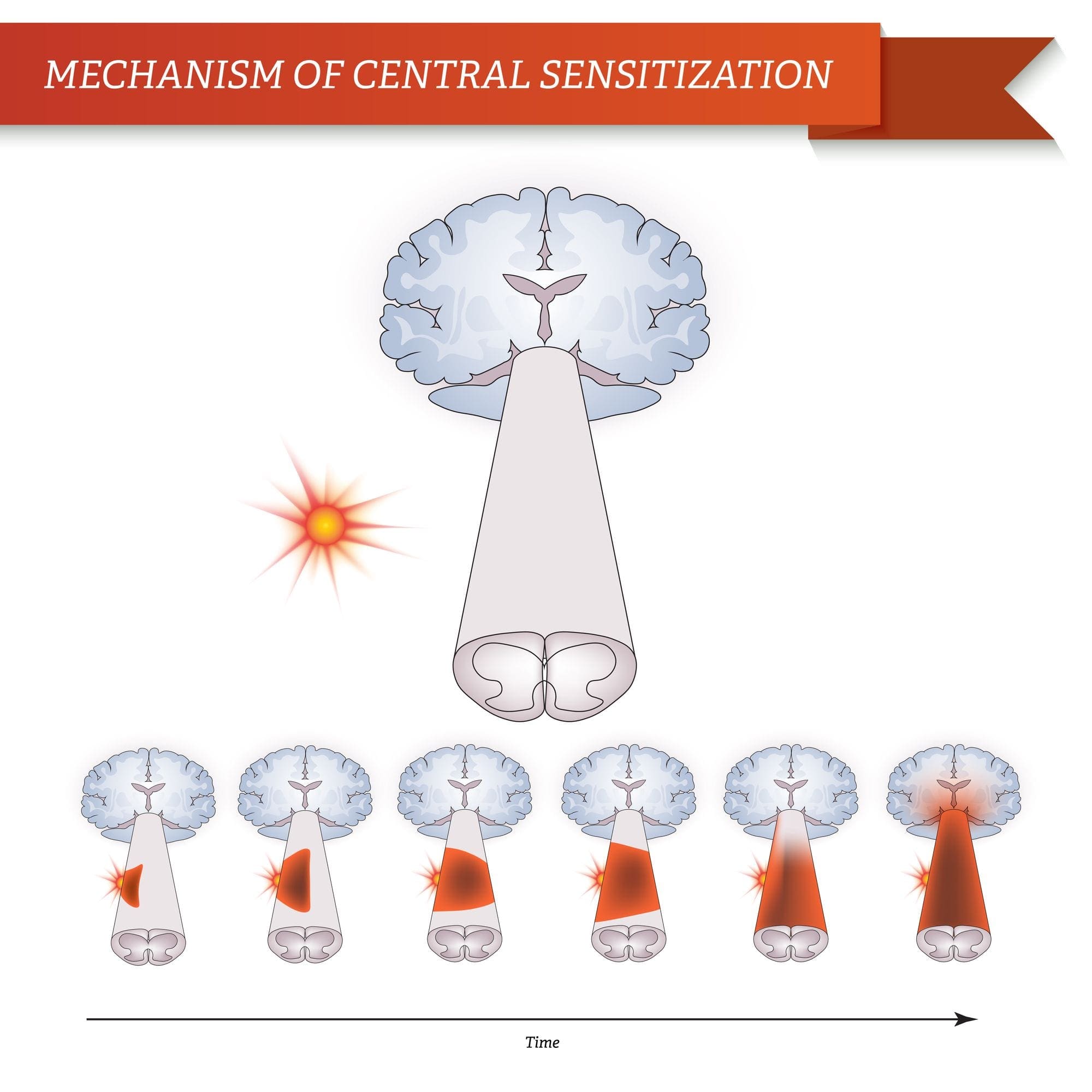
Despois dunha lesión nerviosa periférica, as neuronas fanse máis sensibles e desenvolven excitabilidade anormal e elevada sensibilidade á estimulación.
Isto é coñecido como ... Sensibilización Periférica!
MECANISMOS CENTRALES
Como consecuencia da actividade espontánea en curso na periferia, as neuronas desenvolven unha maior actividade de fondo, campos receptivos ampliados e un aumento nas respostas aos impulsos aferentes, incluíndo estímulos táctiles normais
Isto é coñecido como ...Sensibilización Central!
CAUSAS COMUNES
As lesións ou as enfermidades do sistema nervioso somatosensorial poden provocar unha transmisión alterada e desordenada de sinais sensoriais na medula espiñal e no cerebro; As condicións comúns asociadas á dor neuropática inclúen:
Neuralxia postherpética
Neuralgia trigeminal
Radiculopatía dolorosa
Neuropatia diabética
A infección polo VIH
Lepra
Amputación
Dor periférica nas lesións nerviosas
Accidente cerebrovascular (en forma de dor central post-derrame)
DOR DE EXTREMIDADE FANTÁMICA E REALIDADE AUGMENTADA
Pena de limbo fantasma e AR
INFLAMAMACIÓN NEUROGÉNICA
Obxectivos:
¿Que é iso?
Cal é a fisiopatoloxía detrás dela?
Cales son as causas?
Como podemos solucionalo?
INFLAMAMACIÓN NEUROGÉNICA
Inflamación neurogénica é unha resposta inflamatoria local provocada neuronalmente caracterizada por vasodilatación, aumento da permeabilidade vascular, desgranulación de mastocitos e liberación de neuropéptidos, incluíndo SP e péptido relacionado co xene de calcitonina (CGRP)
Parece que desempeña un papel importante na patoxenia das numerosas enfermidades que inclúen a enxaqueca, psoriase, asma, fibromialxia, eczema, rosácea, distonía e sensibilidade química múltiple.
CAUSAS COMUNES
Existen varios camiños polos que se pode iniciar a inflamación neurogénica. Está ben documentado, utilizando tanto modelos animais como neuronas illadas in vitro, que a capsaicina, a calor, os protóns, a bradiquinina ea tryptase son reguladores de fluxo do influxo de calcio intracelular, o que produce unha liberación inflamatoria de neuropéptidos. En contraste, pénsase que as prostaglandinas E2 e I2, citocinas, interleukina-1, interleukina-6 e factor de necrose tumoral non causan a liberación do neurotransmisor, senón excitar as neuronas sensoriais e, así, reducir o limiar de disparo e causar a liberación aumentada neuropéptidos.
Aínda que a inflamación neurogénica foi ampliamente estudada e ben documentada nos tecidos periféricos, ata hai pouco o concepto de inflamación neurogénica no SNC mantívose en gran medida inexplorado. Dada a capacidade de inflamación neurogénica para influír na permeabilidade vascular e levar á xénese do edema, agora foi ampliamente investigado polo seu potencial para influenciar a permeabilidade do BBB eo edema vasogénico dentro do cerebro e da medula espiñal baixo diferentes condicións patolóxicas.
Todos nos ensinaron como nenos que hai sentidos 5: vista, gusto, son, olor e toque. Os catro sentidos iniciais utilizan órganos claros e distintos, como os ollos, as papilas gustativas, as orellas e o nariz, pero ¿como toca o sentido do corpo exactamente? O toque experimenta todo o corpo, tanto dentro como fóra. Non hai un órgano distinto que se encarga de detectar o toque. Ao contrario, hai diminutos receptores ou terminacións nerviosas ao redor de todo o corpo que senten o toque onde se produce e envía sinais ao cerebro con información sobre o tipo de toque que se produciu. Como un paladar da lingua detecta o sabor, os mecanorreceptores son glándulas dentro da pel e noutros órganos que detectan sensacións de tacto. Coñécense como mecanorreceptores porque están deseñados para detectar sensacións mecánicas ou diferenzas de presión.
Función dos mecanorreceptores
Unha persoa entende que experimentaron unha sensación unha vez que o órgano encargado de descubrir ese sentido específico envía unha mensaxe ao cerebro, que é o órgano principal que procesa e organiza toda a información. As mensaxes envíanse de todas as áreas do corpo ao cerebro a través de fíos referidos como neuronas. Hai miles de pequenas neuronas que se ramifican a todas as áreas do corpo humano e nas finais de moitas destas neuronas están mecanorreceptores. Para demostrar o que pasa cando toca un obxecto, usaremos un exemplo.
Imaxina que un mosquito aterra no brazo. A cepa deste insecto, tan lixeiro, estimula aos mecanorreceptores nesa determinada área do brazo. Estes mecanorreceptores envían unha mensaxe ao longo da neurona á que están conectados. A neurona conecta todo o camiño ata o cerebro, que recibe a mensaxe de que algo está tocando o seu corpo na localización exacta do mecanorreceptor específico que enviou a mensaxe. O cerebro actuará con este consello. Quizais lle diga aos ollos que miran a rexión do brazo que detectou a sinatura. E cando os ollos din o cerebro que hai un mosquito no brazo, o cerebro pode dicirlle á man que o repele rapidamente. Así funcionan os mecanorreceptores. O obxectivo do artigo seguinte é demostrar e discutir en detalle a organización funcional e os determinantes moleculares dos mecanorreceptores.
Táctil: Organización funcional e determinantes moleculares de receptores mecanosensibles
Abstracto
Os mecanorreceptores cutáneos localízanse nas distintas capas da pel onde detectan unha ampla gama de estímulos mecánicos, incluíndo o cepillo lixeiro, o estiramento, a vibración ea presión nociva. Esta variedade de estímulos coincide cunha variedade diversa de mecanorreceptores especializados que responden de forma específica á deformación cutánea e transmiten estes estímulos a estruturas cerebrais máis altas. Os estudos realizados a través de mecanorreceptores e terminacións nerviosas sensoriais tratábeis xeneticamente comezan a descubrir mecanismos de sensación táctil. O traballo neste campo deu aos investigadores un coñecemento máis profundo da organización do circuíto subxacente á percepción do tacto. As novas canles iónicas xurdiron como candidatos para as moléculas de transdución e as propiedades das correntes pechadas mecánicamente melloraron a comprensión dos mecanismos de adaptación a estímulos táctiles. Esta revisión destaca os avances na caracterización das propiedades funcionais dos mecanorreceptores en canles iónicos e peludos e peles que detectan insumos mecánicos e adaptan o mecanógrafo.
Palabras clave:mecanorreceptor, canle mecanosensible, dor, pel, sistema somatosensorial, toque
introdución
O toque é a detección de estímulos mecánicos que afectan a pel, incluídos estímulos mecánicos inócuis e nocivos. É un sentido esencial para a supervivencia e o desenvolvemento de mamíferos e humanos. O contacto de obxectos sólidos e fluídos coa pel dá información necesaria ao sistema nervioso central que permite a exploración e recoñecemento do ambiente e inicia a locomoción ou o movemento planificado da man. O toque tamén é moi importante para a aprendizaxe, os contactos sociais e a sexualidade. O sentido do toque é o sentido menos vulnerable, aínda que pode ser distorsionado (hiperestesia, hipoestesia) en moitas condicións patolóxicas.1-3
As respostas táctiles implican unha codificación moi precisa da información mecánica. Os mecanorreceptores cutáneos localízanse nas distintas capas da pel onde detectan unha ampla gama de estímulos mecánicos, incluíndo cepillo lixeiro, estiramento, vibración, desviación do cabelo e presión nociva. Esta variedade de estímulos é igualada por unha variedade diversa de mecanorreceptores especializados que responden á deformación cutánea dun xeito específico e transmiten estes estímulos a estruturas cerebrais superiores. As neuronas somatosensoriais da pel divídense en dous grupos: os mecanorreceptores de baixo limiar (LTMR) que reaccionan á presión benigna e os mecanorreceptores de alto limiar (HTMR) que responden á estimulación mecánica nociva. Os corpos celulares LTMR e HTMR residen dentro dos ganglios da raíz dorsal (DRG) e dos ganglios sensoriais craniais (ganglios do trixemino). As fibras nerviosas asociadas con LTMR e HTMR clasifícanse como fibras A?-, A?- ou C en función das súas velocidades de condución do potencial de acción. As fibras C non están amielínicas e teñen as velocidades de condución máis lentas (~2 m/s), mentres que A? e A? As fibras están lixeiramente e moi mielínicas, mostrando velocidades de condución intermedias (~12 m/s) e rápidas (~20 m/s), respectivamente. Os LTMR tamén se clasifican como respostas de adaptación lenta ou rápida (SA- e RA-LTMR) segundo as súas taxas de adaptación ao estímulo mecánico sostido. Distínguense ademais polos órganos extremos cutáneos que inervan e os seus estímulos preferidos.
A capacidade dos mecanorreceptores de detectar sinais mecánicos depende da presenza de canles iónicos mecano-transdutores que transforman rapidamente as forzas mecánicas en sinais eléctricos e despolarizan o campo receptivo. Esta despolarización local, chamada potencial de receptor, pode xerar potenciais de acción que se propagan cara ao sistema nervioso central. Non obstante, as propiedades das moléculas que median a mecanotransdución e a adaptación ás forzas mecánicas non están claras.
Nesta revisión, ofrecemos unha visión xeral das propiedades dos mecanorreceptores dos mamíferos no tacto inocuo e nocivo na pel peluda e glabra. Tamén consideramos o coñecemento recente sobre as propiedades das correntes mecánicas para tentar explicar o mecanismo de adaptación dos mecanorreceptores. Finalmente, revisamos os avances recentes realizados na identificación de canles iónicos e proteínas asociadas responsables da xeración de correntes mecano-dependidas.
Toque inocuo
LTMRs asociados ao folículo capilar
Os folículos capilares representan mini-órganos que producen un cabelo que detectan un toque lixeiro. As fibras asociadas con folículos pilosos responden ao movemento do cabelo e á súa dirección disparando trens de potenciais de acción ao inicio e eliminación do estímulo. Están adaptando rapidamente os receptores.
Gato e coello. No pelaje do gato e do coello, os folículos pilosos pódense dividir en tres tipos de folículos pilosos, o pelo Down, o Guard e os Tylotrichs. Os pelos de Down (debaixo do pelo, la, vellus)4 son os máis numerosos, os máis curtos e finos do pelaje. Son ondulados, incoloros e xurdiron en grupos de dous a catro pelos dun orificio común na pel. Os pelos da Garda (monotrichs, overhears, tophair)4 son lixeiramente curvados, pigmentados ou non pigmentados, e xurdiron individualmente das bocas dos seus folículos. Os tilótricos son os menos numerosos, os máis longos e grosos.5,6 Son pigmentados ou non pigmentados, ás veces ambos e xurdiron individualmente dun folículo que está rodeado por un asa de vasos sanguíneos capilares. As fibras sensoriais que fornecen a un folículo piloso sitúase debaixo da glándula sebácea e atribúense a A? ou fibras A?-LTMR.7
En estreita aposición ao eixe do cabelo baixo, xusto debaixo do nivel da glándula sebácea está o anel de finais lanceoladas de pilo-Ruffini. Estas terminacións nerviosas sensoriais sitúanse nun curso espiral ao redor do eixe do cabelo dentro do tecido conxuntivo que forma o folículo piloso. Dentro do folículo piloso, hai tamén terminacións nerviosas libres, algunhas delas formando mecanorreceptores. Frecuentemente, os corpúsculos táctiles (ver a pel glabra) están rodeando a rexión do pescozo do folículo tylotrich.
No período 1930-1970 exploráronse intensamente as propiedades das terminacións nerviosas mielínicas na pel peluda de gato e coello (revisión en Hamann, 1995).8 Sorprendentemente, Brown e Iggo, estudando 772 unidades con fibras nerviosas aferentes mielinizadas dos nervios safenos dos gatos. e coello, clasificaron as respostas en tres tipos de receptores correspondentes aos movementos de cabelos Down (receptores tipo D), cabelos Guard (receptores tipo G) e cabelos Tylotrich (receptores tipo T).9 Reuníronse todas as respostas das fibras nerviosas aferentes. no receptor de adaptación rápida do tipo I (RA I) por oposición ao receptor de Pacini denominado RA II. Os mecanorreceptores RA I detectan a velocidade do estímulo mecánico e teñen un bordo nítido. Non detectan variacións térmicas. Burgess et al. tamén describiu un receptor de campo de rápida adaptación que responde de forma óptima ao acariciamento da pel ou ao movemento de varios pelos, o que se atribuíu á estimulación das terminacións pilo-Ruffini. Ningunha das respostas do folículo piloso foi atribuída á actividade da fibra C.10
Ratos. Na pel pelada dos ratos, descríbense tres tipos principais de folículos pilosos: zigzag (ao redor de 72%), awl / auchene (ao redor de 23%) e garda ou tylotrich (ao redor de 5%). 11-14 Zigzag e Awl / Os folículos de cabelo auchenne producen os cabelos máis delgados e máis curtos e están asociados a unha glándula sebácea. Os pelos de garda ou tylotrich son os máis longos dos folículos pilosos. Caracterízanse por unha gran lámpada capilar asociada a dúas glándulas sebáceas. Os pelos de garda e punzón / aucheno están dispostos nun modelo iterativo, regularmente espazado, mentres que os pelos en zigzag densamente poboan as áreas da pel que rodean os dous tipos de folículo pilosos máis grandes [fig. 1 (A1, A2 e A3)].
Figura 1. Organización e proxección de mecanorreceptores cutáneos. Na pel pelada, o pincel lixeiro e o toque son detectados principalmente pola inervación arredor dos folículos pilosos: awl / auchenne (A1), zigzag (A2) e garda (A3). Os pelos de punzón/auchene están triplemente inervados polas terminacións lanceoladas C-LTMR (A4), A?-LTMR e A? adaptación rápida-LTMR (A6). Os folículos pilosos en zigzag son os tallos máis curtos e están inervados tanto por C-LTMR (A4) como por A? -LTMR terminacións lanceoladas (A5). Os folículos pilosos de garda máis longos están inervados por A? terminacións lanceoladas lonxitudinais de LTMR de adaptación rápida (A6) e están asociadas con A? adaptando lentamente-LTMR das terminacións de cúpula táctil (A7). As proxeccións centrais de todas estas fibras rematan en láminas distintas, pero parcialmente superpostas, do corno dorsal da medula espiñal (C-LTMR na lámina II, A?-LTMR na lámina III e A?-LTMR nas láminas IV e V). As proxeccións de LTMR que inervan o mesmo ou os folículos do pelo adxacentes están aliñados para formar unha columna estreita no corno dorsal da medula espiñal (B1 en gris). Só na pel pelada, unha subpopulação de fibras C sen finalización inerva a epiderme e responde a un toque agradable (A8). Estas fibras C-touch non responden ao tacto nocivo e aínda non se coñece o seu percorrido (B2). Na pel glabra, o toque inocuo está mediado por catro tipos de LTMRs. O complexo celular-neurita de Merkel está na capa basal da epiderme (C1). Este mecanorreceptor consiste nunha disposición entre moitas células de Merkel e unha terminal nerviosa agrandada dunha única A? fibra. As células de Merkel mostran procesos similares aos dedos que contactan con queratinocitos (C2). O final de Ruffini localízase na derme. É unha fina forma de puro encapsulado con terminacións sensoriais conectadas a A? fibra (C3). O corpúsculo de Meissner conectado a A? terminación nerviosa e está situada nas papilas dérmicas. Este mecanorreceptor encapsulado consta de células de apoio envasadas dispostas como lamelas horizontais rodeadas de tecido conxuntivo (C4). O corpúsculo pacinian é o mecanorreceptor máis profundo. Unha única A? a terminación nerviosa non amielínica remata no centro deste gran corpúsculo ovoide formado por lamelas concéntricas. As proxeccións destas fibras A?-LTMR na medula espiñal divídense en dúas ramas. A rama central principal (B3) ascende na medula espiñal nos fascículos cuniléos ou gracios (B5) formando a medula ipsilateral no nivel de medula onde as aferencias primarias fan a súa primeira sinapsis (B6). As neuronas secundarias fan unha decussación sensorial (B7) para formar un tracto no lemnisco medial que ascende polo tronco cerebral ata o cerebro medio, especialmente no tálamo. A branca secundaria do LTMR termina no corno dorsal na lámina II, IV, V e interfire coa transmisión da dor (B4). O toque nocivo é detectado pola terminación do nervio libre na epiderme de pel peludo (A9) e glabro (C7). Estes mecanorreceptores son a terminación de A?-HTMR e C-HTMR en estreito contacto cos queratinocitos veciños (C6). A ?-hTMR termina na lámina I e V; C-HTMR termina na lámina I e II (B8). Ao nivel do corno dorsal da medula espiñal, as afeccións primarias HTMRs fan sinapses con neuronas secundarias que cruzan a liña media e suben á estrutura cerebral máis alta no fascículo anterolateral (B9, B10).
Recentemente, Ginty e os seus colaboradores utilizaron unha combinación de etiquetaxe xenética molecular e enfoques de trazado retrógrado somatotópico para visualizar a organización das terminacións axonales periféricas e centrais dos LTMR en ratos.15 Os seus descubrimentos apoian un modelo no que as características individuais dun estímulo táctil complexo son extraído polos tres tipos de folículo piloso e transmitido a través das actividades de combinacións únicas de fibras A?-, A?- e C- ao corno dorsal.
Mostraron que o etiquetado xenético das neuronas DRG tirosina hidroxilase positiva (TH+) caracteriza unha poboación de neuronas sensoriais de pequeno diámetro e non peptidérxicas e permite a visualización das terminacións periféricas C-LTMR na pel. Sorprendentemente, descubriuse que as ramas axoneais dos C-LTMR individuais se arborizan e forman terminacións lanceoladas lonxitudinais que están íntimamente asociadas con zigzag (80% das terminacións) e punzón / auchene (20% das terminacións), pero non con folículos pilosos tilotrópicos [Fig. 1 (A4)]. Durante moito tempo pensouse que as terminacións lanceoladas lonxitudinais pertencen exclusivamente aos A?-LTMR e, polo tanto, era inesperado que as terminacións dos C-LTMR formasen terminacións lanceoladas lonxitudinais.15 Estes C-LTMR teñen unha adaptación intermedia en comparación cos que se adaptan lenta e rapidamente. mecanorreceptores mielinizados [Fig. 2 (C1)].
Figura 2. Receptores táctiles en mamíferos: os receptores táctiles cutáneos diferéncianse en tacto inocuo apoiados por múltiples receptores con baixo limiar mecánico (LTMR) na pel glabra e peluda e toque nocivo apoiado por receptores de alto limiar mecánico (HTMR). Constitúen terminacións libres nerviosas que terminan principalmente na epiderme. (A) Pel glabra. A1: Os corpúsculos de Meissner detectan o movemento da pel e o deslizamento do obxecto na man. Son importantes para a manipulación de obxectos e a destreza. Os receptores adáptanse rapidamente ao estímulo, están conectados a A? fibras e fibras escasamente a C e teñen un campo receptor grande. A2: Os corpúsculos de Ruffini detectan o estiramento da pel e son importantes para detectar a posición dos dedos e o obxecto de entrega. O receptor adáptase lentamente ao estímulo e mantivo a actividade mentres se aplicase o estímulo. Os receptores están conectados a A? fibras e teñen gran campo receptivo. A3: os corpúsculos de Pacini son máis profundos na derme e detectan vibracións. Os receptores están conectados a A? fibras; adáptanse rapidamente ao estímulo e teñen o campo receptivo máis grande. (B) Pel enteira. B1: Os complexos de células de Merkel están presentes tanto na pel glabra como arredor do cabelo. Exprésanse densamente na man e son importantes para a percepción da textura e a mellor discriminación entre dous puntos. Son responsables da precisión dos dedos. Os receptores están conectados a A? fibras; adáptanse lentamente ao estímulo e teñen un campo receptivo curto. B2: HTMR de tacto nocivo con adaptación moi lenta ao estímulo, é dicir, activos mentres se aplique o estímulo nociceptivo. Están formados pola terminación nerviosa libre de A? e fibras C asociadas aos queratinocitos. (C) Pel peluda. C1: os folículos pilosos están asociados aos diferentes tipos de cabelo. Nos ratos, os pelos de garda son os máis longos e pouco expresados, os punzóns/auchennes son de tamaño mediano e os en zigzag son os máis pequenos e os máis densamente expresados. Están conectados a A? fibras pero tamén a A? e fibras C-LTMRs para cabelos de punzón/auchenne e zizag. Detectan o movemento do cabelo, incluíndo un tacto agradable durante a caricia. Adáptanse rapidamente ou con cinética intermedia ao estímulo. C2: as terminacións nerviosas C-touch corresponden a un subtipo de terminación de fibras C con terminación libre caracterizada por un baixo limiar mecánico. Suponse que codifican para unha sensación agradable inducida pola caricia. Adáptanse moderadamente ao estímulo e teñen un campo receptivo curto. As canles iónicas supostas mecanosensibles (MS) expresadas nos diferentes receptores táctiles indícanse segundo os datos preliminares e resumen a hipótese actual en avaliación.
Unha segunda poboación importante identificada refírese ás terminacións A?-LTMR en Awl/Auchenne e aos folículos en zigzag para comparar co folículo piloso Down moi estudado en gatos e coellos. Ginty e os seus colaboradores demostraron que TrkB se expresa en niveis altos nun subconxunto de neuronas DRG de diámetro medio. As gravacións intracelulares que empregan a preparación ex vivo do nervio da pel de fibras marcadas revelaron que exhiben as propiedades fisiolóxicas das fibras previamente estudadas en gatos e coellos: sensibilidade mecánica exquisita (limiar de Von Frey < 0.07 mN), adaptación rápida das respostas a estímulos supralimiares, condución intermedia. velocidades (5.8 � 0.9 m/s) e espigas de soma non flexionadas estreitas.15 Estes A?-LTMR forman terminacións lanceoladas lonxitudinais asociadas con practicamente todos os folículos pilosos en zigzag e punzón/auchene do tronco [Fig. 1 (A5)].
Finalmente, demostraron que as terminacións periféricas de A? Os LTMR forman terminacións lanceoladas lonxitudinais asociadas con folículos pilosos guard (ou tylotrich) e awl/auchene [Fig. 1 (A6)].15 Ademais, os pelos de Guard tamén están asociados cun complexo celular de Merkel que forma unha cúpula táctil conectada a A? LTMR adaptando lentamente [Fig. 1 (A7)].
En resumo, practicamente todos os folículos pilosos en zigzag están inervados por terminacións lanceoladas tanto C-LTMR como A?-LTMR; os pelos de punzón/auchene están triplemente inervados por A? terminacións lanceoladas-LTMR, A?-LTMR e C-LTMR de adaptación rápida; Os folículos pilosos protectores están inervados por A? terminacións lanceoladas lonxitudinais de LTMR de adaptación rápida e interactúan con A? adaptando lentamente-LTMR de terminacións de cúpula táctil. Así, cada folículo piloso do rato recibe combinacións únicas e invariables de terminacións LTMR correspondentes a órganos finais mecanosensoriais neurofisioloxicamente distintos. Tendo en conta a disposición iterativa destes tres tipos de cabelo, Ginty e os seus colaboradores propoñen que a pel pilosa consiste nunha repetición iterativa da unidade periférica que contén: (1) un ou dous pelos de garda situados no centro, (2) ~20 pelos de punzón/auchenne circundantes e (3). ) ~80 pelos en zigzag intercalados [Fig. 2 (C1)].
Proxección da medula espiñal. As proxeccións centrais de A? Os LTMR, A?-LTMR e C-LTMR de adaptación rápida terminan en láminas distintas, pero parcialmente superpostas (II, III, IV) do corno dorsal da medula espiñal. Ademais, os terminais centrais dos LTMR que inervan os mesmos folículos pilosos ou adxacentes dentro dunha unidade LTMR periférica están aliñados para formar unha estreita columna LTMR no corno dorsal da medula espiñal [Fig. 1 (B1)]. Así, parece probable que unha cuña ou columna de terminacións aferentes sensoriais primarias organizadas somatotópicamente no corno dorsal represente o aliñamento das proxeccións centrais de A?-, A?- e C-LTMR que inervan a mesma unidade periférica e detectan estímulos que actúan sobre o mesmo pequeno grupo de folículos pilosos. Baseándose no número de pelos de garda, punzón/auchene e en zigzag do tronco e das extremidades e os números de cada subtipo LTMR, Ginty e os seus colaboradores estiman que o corno dorsal do rato contén 2,000-4,000 columnas LTMR, o que corresponde ao número aproximado de periféricos. Unidades LTMR.15
Ademais, os axóns dos subtipos LTMR están estreitamente asociados entre si, tendo proxeccións entrelazadas e terminacións lanceoladas interdixitadas que inervan o mesmo folículo piloso. Ademais, debido a que os tres tipos de folículos pilosos presentan diferentes formas, tamaños e composicións celulares, é probable que teñan distintas propiedades de axuste de deflexión ou vibración. Estes achados son consistentes coas medicións neurofisiolóxicas clásicas no gato e coello que indican que A? Os RA-LTMR e A?-LTMR pódense activar de forma diferencial pola desviación de distintos tipos de folículo piloso.16,17.
En conclusión, o toque na pel pelada é a combinación de: (1) os números relativos, distribucións espaciais únicas e distintas propiedades morfolóxicas e deflexivas dos tres tipos de folículos pilosos; (2) as combinacións únicas de finais de subtipos LTMR asociados a cada un dos tres tipos de folículos pilosos; e (3) sensibilidades distintas, velocidades de condución, patróns de tren de espiga e propiedades de adaptación das catro clases principais de LTMRs asociados ao folículo foliar que permiten ao sistema peles peludo mecanosensorial extraer e transmitir ao SNC as combinacións complexas de calidades que definen a tocar.
LTMRs de fin de nervios libres
Xeralmente, as terminacións libres de fibras C na pel son HTMR, pero unha subpoboación de fibras C non responde ao tacto nocivo. Este subconxunto de aferentes táctiles de fibra C (CT) representa un tipo distinto de unidades mecanorreceptivas de baixo limiar e non amielínicas existentes na pel peluda pero non glabra de humanos e mamíferos [Fig. 1 (A8)].18,19 As TC asócianse xeralmente coa percepción dunha estimulación táctil agradable no contacto corporal.20,21
As aferentes CT responden a forzas de indentación no intervalo de 0.3-2.5 mN e son, polo tanto, tan sensibles á deformación da pel como moitos dos A? aferentes.19 As características de adaptación das aferentes TC son, polo tanto, intermedias en comparación cos mecanorreceptores mielínicos de adaptación lenta e rápida. Os campos receptivos das aferentes da TC humana son de forma aproximadamente redonda ou oval. O campo consta de un a nove pequenos puntos sensibles distribuídos nunha área de ata 35 mm2.22 Os receptores homólogos do rato están organizados nun patrón de parches descontinuos que cobren preto do 50-60% da área da pel peluda [Fig. 2 (C2)].23
As evidencias de pacientes sen afeccións táctiles mielinizadas indican que a sinalización das fibras de CT activa o córtex insular. Dado que este sistema é pobre en codificar aspectos discriminatorios do toque, pero ben adaptado para codificar un toque lento e suave, as fibras de CT na pel pel poden formar parte dun sistema para procesar aspectos agradables e socialmente relevantes do toque. teñen un papel na inhibición da dor e propúxose recentemente que a inflamación ou o trauma poden cambiar a sensación transmitida polos LTMRs de fibra C de toque agradable a dor.
Que vías de afección de TC de vía aínda non se sabe [fig. 1 (B2)], pero documentáronse as entradas táctiles de baixo límite para as células de proxección espinotámica, dándolle credibilidade a informes de débiles sutís e contralaterales de detección de toque en pacientes humanos tras a destrución destas vías despois dos procedementos de cordotomia.
LTMRs en pel Glabrous
Complexos celular-neurita de Merkel e cúpula táctil. Merkel (1875) foi a primeira en dar unha descrición histolóxica de grupos de células epidérmicas con grandes núcleos lobulados, que entran en contacto con presuntas fibras nerviosas aferentes. Asumiu que serviu o sentido do tacto chamándoas Tastzellen (células táctiles). Nos humanos, os complexos de neuritas de células de Merkel están enriquecidos en áreas sensibles ao tacto da pel, atópanse na capa basal da epiderme dos dedos, os beizos e os xenitais. Tamén existen na pel peluda de menor densidade. O complexo de neuritas da célula de Merkel consiste nunha célula de Merkel en estreita aposición a unha terminal nerviosa agrandada a partir dunha única célula mielínica A? fibra [Fig. 1 (C1)] (revisión en Halata e colaboradores).29 No lado epidérmico a célula de Merkel presenta procesos similares a dedos que se estenden entre os queratinocitos veciños [Fig. 1 (C2)]. As células de Merkel son células epidérmicas derivadas de queratinocitos.30,31 O termo de cúpula táctil foi introducido para denominar a gran concentración de complexos de células de Merkel na pel peluda da pata dianteira do gato. Unha cúpula táctil podería ter ata 150 células de Merkel inervadas por unha soa fibra A? e nos humanos ademais das fibras A?, A? e as fibras C tamén estaban presentes regularmente.32-34
A estimulación dos complexos de neuritas de células de Merkel dá lugar a respostas de tipo I (SA I) de adaptación lenta, que se orixinan a partir de campos receptivos puntuados con bordos nítidos. Non hai descarga espontánea. Estes complexos responden á profundidade de indentación da pel e teñen a maior resolución espacial (0.5 mm) dos mecanorreceptores cutáneos. Transmiten unha imaxe espacial precisa dos estímulos táctiles e proponse que son os responsables da discriminación de forma e textura [Fig. 2 (B1)]. Os ratos carentes de células de Merkel non poden detectar superficies texturizadas cos seus pés mentres o fan usando os seus bigotes.35
Se a célula de Merkel, a neurona sensorial ou ambas son sitios de mecanotransdución aínda é un tema de debate. Nas ratas, a destrución fototóxica das células de Merkel elimina a resposta SA I.36 En ratos con células de Merkel suprimidas xeneticamente, a resposta SA I rexistrada na preparación ex vivo da pel/nervio desapareceu por completo, demostrando que as células de Merkel son necesarias para a correcta codificación de Merkel. respostas dos receptores.37 Non obstante, a estimulación mecánica de células de Merkel illadas en cultivo por presión impulsada por motor non xera correntes mecánicas.38,39 Os queratinocitos poden desempeñar un papel importante no funcionamento normal do complexo neurítico da célula de Merkel. Os procesos similares aos dedos das células de Merkel poden moverse coa deformación da pel e o movemento das células da epiderme, e este pode ser o primeiro paso da transdución mecánica. Claramente, as condicións necesarias para estudar a mecanosensibilidade das células de Merkel aínda non se estableceron.
Remates de Ruffini. As terminacións Ruffini son finas terminacións sensoriais encapsuladas en forma de puro conectadas a A? terminacións nerviosas. As terminacións Ruffini son pequenos cilindros de tecido conxuntivo dispostos ao longo de fíos de coláxeno dérmico que son subministrados por unha a tres fibras nerviosas mielínicas de 4 µm de diámetro. Ata tres cilindros de diferente orientación na derme poden fusionarse para formar un receptor [Fig. 6 (C1)]. Estruturalmente, as terminacións de Ruffini son similares aos órganos do tendón de Golgi. Exprésanse amplamente na derme e identificáronse como os mecanorreceptores cutáneos tipo II (SA II) de adaptación lenta. No contexto da actividade nerviosa espontánea, unha descarga regular de adaptación lenta prodúcese por estimulación mecánica mantida con forza baixa perpendicular ou máis eficazmente por estiramento dérmico. A resposta SA II orixínase de grandes campos receptivos con fronteiras escuras. Os receptores de Ruffini contribúen á percepción da dirección do movemento do obxecto a través do patrón de estiramento da pel [Fig. 3 (A2)].
Nos ratos, as respostas SA I e SA II pódense separar electrofisioloxicamente na preparación ex-vivo da pel nerviosa. 40 Nandasena e colaboradores informaron sobre a inmunolocalización da aquaporina 1 (AQP1) nas terminacións periodontais de Ruffini dos incisivos de rato suxerindo que AQP1 está implicado en o mantemento do equilibrio osmótico dental necesario para a mecanotransdución. 41 As terminacións periodontais Ruffini tamén expresaron o suposto canle de ións mecanosensible ASIC3.42
Corpúsculos de Meissner. Os corpúsculos de Meissner localízanse nas papilas dérmicas da pel glabra, principalmente nas palmas das mans e nas plantas dos pés, pero tamén nos beizos, na lingua, na cara, nos pezones e nos xenitais. Anatomicamente, consisten nunha terminación nerviosa encapsulada, a cápsula está feita de células de apoio aplanadas dispostas como lamelas horizontais incrustadas no tecido conxuntivo. Hai unha única fibra nerviosa A? aferentes conectadas por corpúsculo [Fig. 1 (C4)]. Calquera deformación física do corpúsculo desencadea unha descarga de potenciais de acción que cesa rapidamente, é dicir, están a adaptarse rapidamente aos receptores. Cando se elimina o estímulo, o corpúsculo recupera a súa forma e ao facelo produce outra descarga de potenciais de acción. Debido á súa localización superficial na derme, estes corpúsculos responden selectivamente ao movemento da pel, á detección táctil do escorregamento e ás vibracións (20-40 Hz). Son sensibles á pel dinámica, por exemplo, entre a pel e un obxecto que se está a manipular [Fig. 2 (A1)].
Corpúsculos pacinianos. Os corpúsculos de Pacini son os mecanorreceptores máis profundos da pel e son os mecanorreceptores cutáneos encapsulados máis sensibles do movemento da pel. Estes grandes corpúsculos ovoides (1 mm de lonxitude) feitos de láminas concéntricas de tecido conxuntivo fibroso e fibroblastos revestidos por células de Schwann modificadas planas exprésanse na derme profunda.43 No centro do corpúsculo, nunha cavidade chea de líquido chamada bulbo interno. , remata unha única A? terminación nerviosa amielínica aferente [Fig. 1 (C5)]. Teñen un gran campo receptivo na superficie da pel cun centro especialmente sensible. O desenvolvemento e a función de varios tipos de mecanorreceptores de adaptación rápida vense alterados nos ratos mutantes c-Maf. En particular, os corpúsculos de Pacini están gravemente atrofiados.44
Os corpúsculos de Pacini presentan unha adaptación moi rápida en resposta á indentación da pel, a descarga nerviosa II (RA II) de adaptación rápida que son capaces de seguir estímulos vibratorios de alta frecuencia e permiten a percepción de eventos distantes a través de vibracións transmitidas.45 O corpúsculo de Pacinio. as aferentes responden á sangría sostida con actividade transitoria ao inicio e ao desplazamento do estímulo. Tamén se denominan detectores de aceleración porque poden detectar cambios na forza do estímulo e, se se altera a velocidade de cambio do estímulo (como ocorre nas vibracións), a súa resposta faise proporcional a este cambio. Os corpúsculos de Pacini perciben grandes cambios de presión e sobre todo vibracións (150-300 Hz), que poden detectar incluso a centímetros de distancia [Fig. 2 (A3)].
A resposta tónica foi observada no corpuscle pacinian descapsulado. 46 Ademais, os corpúsculos de Pacinian intactos responden con actividade sostida durante estímulos de indentación constante, sen alterar os limiares mecánicos ou a frecuencia de resposta cando a sinalización mediada por GABA está bloqueada entre a glia lamelada e un final nervioso. os compoñentes non neuronais do corpúsculo de Pacinian poden ter un papel dobre no filtrado do estímulo mecánico e na modulación das propiedades de resposta da neurona sensorial.
Proxeccións da medula espiñal. As proxeccións dos A?-LTMR na medula espiñal divídense en dúas ramas. A rama central principal ascende na medula espiñal nas columnas dorsais ipsilaterais ata o nivel cervical [Fig. 1 (B3)]. As ramas secundarias terminan no corno dorsal das láminas IV e interfiren coa transmisión da dor, por exemplo. Isto pode atenuar a dor como parte do control da porta [Fig. 1 (B4)].48
Nos niveis cervicais, os axóns da rama principal sepáranse en dous tractos: o tramo da liña media comprende o fascículo gracioso que transporta información da metade inferior do corpo (pernas e tronco), eo tramo externo comprende o fascículo cuneato que transporta información da metade superior do corpo (brazos e tronco) [fig. 1 (B5)].
As aferentes táctiles primarias fan a súa primeira sinapse con neuronas de segunda orde na médula onde as fibras de cada tracto fan sinapsis nun núcleo do mesmo nome: os axóns do fascículo grácil fan sinapsis no núcleo grácil e os axóns cuneados fan sinapsis no núcleo cuneado [Fig. 1 (B6)]. As neuronas que reciben a sinapse proporcionan as aferentes secundarias e cruzan a liña media inmediatamente para formar un tracto no lado contralateral do tronco encefálico, o lemnisco medial, que ascende a través do tronco encefálico ata a seguinte estación de relevo no mesencéfalo, concretamente no tálamo [Fig. . 1 (B7)].
Especificación molecular de LTMRs. Recentemente elucidaronse parcialmente os mecanismos moleculares que controlan a diversificación precoz dos LTMR. Bourane e colaboradores demostraron que as poboacións neuronais que expresan o receptor Ret tirosina quinase (Ret) e o seu co-receptor GFR?2 en ratos embrionarios E11-13 DRG coexpresan selectivamente o factor de transcrición Mafa.49,50 Estes autores demostran que o Mafa/ As neuronas Ret/GFR?2 destinadas a converterse en tres tipos específicos de LTRM ao nacer: as neuronas SA1 que inervan os complexos de células de Merkel, as neuronas de rápida adaptación que inervan os corpúsculos de Meissner e as aferentes de rápida adaptación (RA I) que forman terminacións lanceoladas arredor dos folículos pilosos. Ginty e os seus colaboradores tamén informan de que as neuronas DRG que expresan Earl-Ret están adaptando rapidamente os mecanorreceptores dos corpúsculos de Meissner, os corpúsculos de Pacini e as terminacións lanceoladas arredor dos folículos pilosos. proxeccións axonales neuronais dentro do tronco cerebral.
Exploración dos mecanorreceptores da pel humana. A técnica de "microneurografía" descrita por Hagbarth e Vallbo en 1968 aplicouse para estudar o comportamento de descarga de terminacións mecanosensibles humanas únicas que abastecen músculo, articulación e pel (ver Macefield, 2005 para a súa revisión).52,53 A maioría da microneurografía da pel humana. os estudos caracterizaron a fisioloxía das aferentes táctiles na pel glabra da man. As gravacións de microelectrodos dos nervios mediano e cubital en suxeitos humanos revelaron a sensación táctil xerada polas catro clases de LTMR: as aferentes de Meissner son particularmente sensibles ao acariciado lixeiro na pel, respondendo ás forzas de cizallamento locais e aos deslizamentos incipientes ou evidentes dentro do campo receptivo. As aferentes pacinianas son exquisitamente sensibles aos transitorios mecánicos rápidos. As aferentes responden con forza ao soprar sobre o campo receptivo. Un corpúsculo de Pacini situado nun díxito adoita responder ao tocar a mesa que apoia o brazo. As aferentes de Merkel teñen característicamente unha alta sensibilidade dinámica aos estímulos de indentación aplicados a unha área discreta e moitas veces responden cunha descarga de descarga durante a liberación. Aínda que as aferentes de Ruffini responden ás forzas aplicadas normalmente á pel, unha característica única das aferentes SA II é a súa capacidade de responder tamén ao estiramento lateral da pel. Finalmente, as unidades capilares do antebrazo teñen grandes campos receptivos ovoides ou irregulares compostos por múltiples puntos sensibles que se correspondían con cabelos individuais (cada unha subministración aferente ~20 pelos).
Sensibilidade mecánica dos queratinocitos
Calquera estímulo mecánico na pel debe transmitirse a través de queratinocitos que forman a epiderme. Estas células omnipresentes poden realizar funcións de sinalización ademais das súas funcións de apoio ou protección. Por exemplo, os queratinocitos segregan ATP, unha importante molécula de sinalización sensorial, en resposta a estímulos mecánicos e osmóticos. 54,55 A liberación de ATP induce un aumento intracelular do calcio por estimulación autocrina de receptores purinérxicos. 55 Ademais, hai evidencias de que a hipotonicidade activa a Rho-quinasa. vía de sinalización e posterior formación de fibras de estrés de actina F que suxire que a deformación mecánica dos queratinocitos pode interferir mecánicamente coas células veciñas como as células de Merkel para o toque inócuo e finais libres de fibras de C para o toque nocivo [fig. 1 (C6)]
Toque nocivo
Os mecanorreceptores de limiar alto (HTMR) son epidérmicos C- e A? terminacións nerviosas libres. Non están asociados a estruturas especializadas e obsérvanse tanto na pel peluda [Fig. 1 (A9)] e pel glabra [Fig. 1(C7)]. Non obstante, o termo de terminación nerviosa libre debe considerarse con prudencia xa que as terminacións nerviosas están sempre en estreita aposición cos queratinocitos ou células de Langherans ou melanocitos. A análise ultraestrutural das terminacións nerviosas revela a presenza de retículo endoplasmático rugoso, abundantes mitocondrias e vesículas de núcleo denso. As membranas adxacentes das células epidérmicas están engrosadas e semellan a membrana postsináptica nos tecidos nerviosos. Teña en conta que as interaccións entre as terminacións nerviosas e as células epidérmicas poden ser bidireccionais xa que as células epidérmicas poden liberar mediadores como ATP, interleucina (IL6, IL10) e bradicinina e, pola contra, as terminacións nerviosas peptidérxicas poden liberar péptidos como CGRP ou substancia P que actúan sobre as células epidérmicas. Os HTMR comprenden mecano-nociceptores excitados só por estímulos mecánicos nocivos e nociceptores polimodais que tamén responden á calor nociva e á substancia química esóxena [Fig. 2 (B2)].58
As fibras aferentes HTMR terminan nas neuronas de proxección no corno dorsal da medula espiñal. Os A?-HTMR entran en contacto con neuronas de segunda orde predominantemente na lámina I e V, mentres que os C-HTMR terminan na lámina II [Fig. 1 (B8)]. As neuronas nociceptivas de segunda orde proxéctanse ao lado controlateral da medula espiñal e ascenden na substancia branca, formando o sistema anterolateral. Estas neuronas terminan principalmente no tálamo [Fig. 1 (B9 e B10)].
Mecano-corrente en neuronas somatosensoriais
Os mecanismos de adaptación lenta ou rápida dos mecanorreceptores aínda non son aclarados. Non está claro en que medida a adaptación do mecanorreceptor é proporcionada polo ambiente celular da terminación dos nervios sensoriais, as propiedades intrínsecas dos canles pechados mecánicamente e as propiedades dos canles iónicos por tensión axonal nas neuronas sensoriais (Fig. 2). Non obstante, os avances recentes na caracterización das correntes pechadas mecánicamente demostraron que existen diferentes clases de canles mecanosensibles nas neuronas DRG e poden explicar algúns aspectos da adaptación dos mecanorreceptores.
O rexistro in vitro en roedores demostrou que o soma das neuronas DRG é intrínsecamente mecanosensible e expresa correntes catiónicas mecano-dependientes.59-64 O vermello gadolinio e rutenio bloquean totalmente as correntes mecanosensibles, mentres que o calcio e o magnesio externos, a concentracións fisiolóxicas, así como a amilorida. e benzamil, provocan bloqueo parcial.60,62,63 FM1-43 actúa como un bloqueador duradeiro, e a inxección de FM1-43 na pata traseira dos ratos diminúe a sensibilidade á dor na proba de Randall�Selitto e aumenta o limiar de retirada da pata avaliado. con pelos de von Frey.65
En resposta á estimulación mecánica sostida, as correntes mecanosensibles diminúen polo peche. En función das constantes de tempo da desintegración da corrente, distinguíronse catro tipos distintos de correntes mecanosensibles: correntes de adaptación rápida (~3 ms), correntes de adaptación intermedia (~6 ms), correntes de adaptación lenta (~15 ms). ) e correntes de adaptación ultralenta (~30 ms).200 Todas estas correntes están presentes con incidencia variable nas neuronas DRG de rata que inervan a pel glabra da pata traseira.300
A sensibilidade mecánica das correntes mecanosensibles pódese determinar aplicando unha serie de estímulos mecánicos incrementais, que permiten unha análise estímulo-corrente relativamente detallada.66 A relación estímulo-corrente é normalmente sigmoidal, e a amplitude máxima da corrente está determinada polo número de canles que están abertas simultáneamente.64,67. Curiosamente, a corrente mecanosensible que se adapta rapidamente mostra un limiar mecánico baixo e un punto medio de media activación en comparación coa corrente mecanosensible de adaptación ultralenta.63,65.
Neuronas sensoriais con fenotipos non nociceptivos que expresan preferentemente correntes mecanosensibles de rápida adaptación cun mínimo limiar mecánico. 60,61,63,64,68 Ás veces, as correntes mecánicas sensibles que se adaptan lentamente e ultra-lentamente son reportadas en supostas células non nociceptivas.64,68 Isto suxeriu que estas correntes contribúan á diferentes limiares mecánicos vistos en LTMRs e HTMRs in vivo. Aínda que estes experimentos in vitro deben ser tomados con cautela, o apoio para a presenza no soma das neuronas DRG de mecano-transdutores de baixo e alto límite tamén foi proporcionado por estimulación baseada en estiramento radial de neuronas sensoriais de rato cultivadas.69 Este paradigma revelou dous principais poboacións de neuronas sensibles ao estiramento, unha que responde a unha amplitude de estímulo baixo e outra que responde selectivamente á amplitude de estímulos elevados.
Estes resultados teñen implicacións importantes, aínda que especulativas, mecanicistas: o limiar mecánico das neuronas sensoriais pode ter pouco que ver coa organización celular do mecanorreceptor pero pode estar nas propiedades dos canles iónicos pechados mecánicamente.
Recentemente desveláronse os mecanismos que subxacen á desensibilización das correntes catiónicas mecanosensibles nas neuronas DRG de rata.64,67 Resulta de dous mecanismos concorrentes que afectan ás propiedades da canle: a adaptación e a inactivación. A adaptación informouse por primeira vez en estudos sobre células ciliadas auditivas. Pódese describir operativamente como unha simple tradución da curva de activación da canle do transductor ao longo do eixe do estímulo mecánico.70-72 A adaptación permite que os receptores sensoriais manteñan a súa sensibilidade a novos estímulos en presenza dun estímulo existente. Non obstante, unha fracción substancial das correntes mecanosensibles nas neuronas DRG non se pode reactivar despois da estimulación mecánica condicionante, o que indica a inactivación dalgunhas canles transdutores.64,67 Polo tanto, tanto a inactivación como a adaptación actúan en conxunto para regular as correntes mecanosensibles. Estes dous mecanismos son comúns a todas as correntes mecanosensibles identificadas nas neuronas DRG de rata, o que suxire que os elementos físico-químicos relacionados determinan a cinética destas canles.64
En conclusión, determinar as propiedades das correntes mecanosensibles endóxenas in vitro é fundamental na procura de identificar mecanismos de transdución a nivel molecular. A variabilidade observada no limiar mecánico e a cinética de adaptación das diferentes correntes de regulación mecánica nas neuronas DRG suxiren que as propiedades intrínsecas das canles iónicas poden explicar, polo menos en parte, o limiar mecánico e a cinética de adaptación dos mecanorreceptores descritos nas décadas de 1960� 80 utilizando preparados ex vivo.
Proteínas mecanosensibles Putativas
As correntes iónicas mecanosensibles nas neuronas somatosensorial están ben caracterizadas, pola contra, pouco se sabe sobre a identidade das moléculas que median a mecanotransdución nos mamíferos. As pantallas xenéticas en Drosophila e C. elegans identificaron moléculas de mecanotransdución candidatas, incluíndo as familias TRP e degenerina / canle Na + (Deg / ENaC). 73 Os intentos recentes para dilucidar as bases moleculares da mecanotransdución nos mamíferos centráronse en gran parte nos homólogos destes candidatos. . Ademais, moitos destes candidatos están presentes en mecanorreceptores cutáneos e neuronas somatosensoriais (Fig. 2).
Canles iónicos de detección de ácido
Os ASIC pertencen a un subgrupo dependente de protóns da familia de canles de Na+ dexenerina epitelial.74 Tres membros da familia ASIC (ASIC1, ASIC2 e ASIC3) exprésanse en mecanorreceptores e nociceptores. O papel das canles ASIC foi investigado en estudos de comportamento utilizando ratos con deleción dirixida de xenes da canle ASIC. A eliminación de ASIC1 non altera a función dos mecanorreceptores cutáneos, senón que aumenta a sensibilidade mecánica das aferentes que inervan o intestino.75 Os ratos knockout ASIC2 presentan unha sensibilidade diminuída dos LTMR cutáneos de adaptación rápida.76 Non obstante, estudos posteriores informaron dunha falta de efectos de eliminación de ASIC2. tanto a mecano-nocicepción visceral como a mecanosensación cutánea.77 A interrupción de ASIC3 diminúe a mecanosensibilidade das aferentes viscerais e reduce as respostas dos HTMR cutáneos aos estímulos nocivos.76
A canle do receptor transitorio
A superfamilia TRP está subdividida en seis subfamilias en mamíferos.78 Case todas as subfamilias TRP teñen membros ligados á mecanosensación nunha variedade de sistemas celulares. 79 En neuronas sensoriais de mamíferos, con todo, os canles TRP son máis coñecidos por detectar información térmica e medir a inflamación neurogénica. e só dúas canles TRP, TRPV4 e TRPA1, foron implicadas na capacidade de resposta. Interrompendo a expresión TRPV4 en ratos ten só efectos modestos nos limiares mecanosensuais agudas, pero reduce fortemente a sensibilidade a estímulos mecánicos nocivos. 80,81 TRPV4 é un determinante crucial na formación das neuronas nociceptivas ao estrés osmótico e á hiperalgesia mecánica durante a inflamación. teñen un papel na hiperalgesia mecánica. Os ratos con deficiencia de TRPA82,83 presentan hipersensibilidade á dor. TRPA1 contribúe á transducción de estímulos mecánicos, fríos e químicos en neuronas sensoriais nociceptor pero parece que non é esencial para a transdución de células capilares.
Non hai ningunha evidencia clara que indica que as canles de TRP e as canles de ASIC expresadas en mamíferos están pechadas mecánicamente. Ningunha destas canles expresadas heteroloxicamente recapitula a sinatura eléctrica das correntes mecanosensibles observadas no seu ambiente nativo. Isto non exclúe a posibilidade de que os canles de ASIC e TRP sexan mecanotransductores, dada a incerteza de se un canal de mecanotransdución pode funcionar fóra do seu contexto celular (ver sección sobre SLP3).
Proteínas Piezo
Os protéicos piezoicos identificáronse recentemente como candidatos prometedores para proteínas mecanosensantes por Coste e os colaboradores. Os vertebrados 86,87 teñen dous membros piezoeléctricos, o Piezo 1 e o Piezo 2, coñecidos anteriormente como FAM38A e FAM38B, respectivamente, que están ben conservados en eucariotas. O piezo 2 é abundante en DRGs, mentres que o Piezo 1 é apenas detectable. Evítanse as correntes mecanosensibles inducidas por piezoi por gadolinio, vermello rutenio e GsMTx4 (unha toxina da tarántula Grammostola spatulata) .88 A expresión de Piezo 1 ou Piezo 2 en sistemas heterólogos produce correntes mecano-sensibles, a cinética de inactivación da corrente Piezo 2 é máis rápida que Piezo 1. Similar ás correntes mecanosensibles endóxenas, as correntes dependentes do piezo teñen potenciais de reversión en torno a 0 mV e non son catiais selectivos, con Na +, K +, Ca2 + e Mg2 + todos permeando a canle subxacente. Do mesmo xeito, as correntes piezo-dependentes están reguladas polo potencial de membrana, cunha marcada desaceleración da cinética actual en potenciais despolarizados.
As proteínas piezoeicas son, sen dúbida, proteínas mecanosensas e comparten moitas propiedades de adaptación rápida de correntes mecanosensibles nas neuronas sensoriais. O tratamento das neuronas cultivadas con DRG con ARN interferente curto 2 piezoeléctrico diminuíu a proporción de neuronas con corrente de adaptación rápida e diminuíu a porcentaxe de neuronas mecanosensibles. Os dominios transmembrana de 86 están localizados en todas as proteínas piezoeléctricas, pero non se atoparon motivos ou cadeas iónicas obvias. identificado. Non obstante, a proteína Piezo 1 do rato se purificou e reconstituíu en bicapas lipídicas asimétricas e forma liposomas canles iónicos sensibles ao vermello rutenio. 87 Un paso esencial na validación da mecanotransdución a través de canles piezoais é empregar enfoques in vivo para determinar a importancia funcional na sinalización táctil. A información foi dada en Drosophila onde a supresión do membro piezo único reduciu a resposta mecánica aos estímulos nocivos, sen afectar o toque normal. 89 Aínda que a súa estrutura segue a ser determinada, esta nova familia de proteínas mecanosensibles é un asunto prometedor para futuras investigacións, máis aló da fronteira de sensación táctil. Por exemplo, un estudo recente sobre pacientes con anemia (xerocitose hereditaria) mostra o papel do piezo 1 no mantemento do volume de eritrocitos homeostasis.90
Transmembrane Channel-Like (TMC)
Un estudo recente indica que dúas proteínas, TMC1 e TMC2, son necesarias para o mecanotransdución de células capilares. 91 A xordeira hereditaria debida á mutación do xene TMC1 foi informada en humanos e ratos. A presenza destes canles aínda non se mostrou no sistema somatosensorial. parece ser un bo avance para investigar.
Proteína similar á estomatina 3 (SLP3)
Ademais das canles de transdución, algunhas proteínas accesorias ligadas á canle demostraron que desempeñan un papel no sentido de tocar a sensibilidade. SLP3 exprésase en neuronas DRG de mamíferos. Os estudos que empregaron ratones mutantes sen SLP3 mostraron un cambio na mecanosensación e correntes mecanosentivas. A función precisa de NXXX SLP94,95 permanece descoñecida. Pode ser un enlazador entre a canle mecanosensible e os microtúbulos subxacentes, tal e como se propón para o seu homólogo C. elegans MEC3 Recentemente GR. O laboratorio de Lewin suxeriu que unha corda é sintetizada por neuronas sensoriais DRG e enlaza unha canle iónica mecanosensible á matriz extracelular. 2.96 Interromper o enlace suprime a corrente RA-mecanosensible suxerindo que algúns canles iónicos son mecanosensibles só cando están ligados. As correntes RA-mecanosensibles tamén son inhibidas pola laminina-97, unha proteína matricial producida polos queratinocitos, reforzando a hipótese dunha modulación da corrente mecanosensitiva por proteínas extracelulares.
K + Subfamilia do canal
Paralelamente ás correntes mecano-sensibles de despolarización catiónica, está a ser investigada a presenza de correntes K + de mecanossensibilidade repolarizantes. Os canais K + nas células mecanosensibles poden entrar no equilibrio actual e contribuír a definir o limiar mecánico e o tempo de adaptación dos mecanorreceptores.
Os membros de KCNK pertencen á familia de canle K + (K2P) do dominio de dous poros. 99,100 O K2P mostra unha notable gama de regulación por axentes celulares, físicos e farmacolóxicos, incluíndo cambios de pH, calor, estiramento e deformación da membrana. Estes K2P están activos no potencial de membrana en repouso. Varias subunidades KCNK están expresadas en neuronas somatosensoriais. As canles 101 KCNK2 (TREK-1), KCNK4 (TRAAK) e TREK-2 están entre os poucos canles para os que se mostrou unha apertura mecánica directa por estiramento de membrana.
Os ratos cun xene KCNK2 alterado mostraron unha maior sensibilidade á calor e estímulos mecánicos leves, pero un limiar de retirada normal para a presión mecánica nociva aplicada á pata traseira mediante a proba de Randall-Selitto. condicións. Os ratos knockout KCNK104 eran hipersensibles á estimulación mecánica leve, e esta hipersensibilidade aumentou coa inactivación adicional de KCNK2 O aumento da mecanosensibilidade destes ratos knockout podería significar que o estiramento normalmente activa as correntes mecanosensibles despolarizantes e repolarizantes de forma coordinada, de forma similar ao desequilibrio de correntes. despolarizando e repolarizando correntes dependentes de voltaxe.
KCNK18 (TRESK) é un dos principais contribuíntes á condutancia de K+ de fondo que regula o potencial de membrana en repouso das neuronas somatosensoriales.106 Aínda que non se sabe se KCNK18 é directamente sensible á estimulación mecánica, pode desempeñar un papel na mediación das respostas ao tacto leve. así como estímulos mecánicos dolorosos. Proponse que KCNK18 e, en menor medida, KCNK3, sexan a diana molecular do hidroxi-?-sanshool, un composto que se atopa nos grans de pementa Schezuan que activa os receptores táctiles e induce unha sensación de formigueo nos humanos.107,108
A canle K + KCNQ4 (Kv7.4) dependente da tensión é crucial para establecer a preferencia de velocidade e frecuencia dunha subpopulação de mecanorreceptores de adaptación rápida tanto en ratos como en humanos. A mutación de KCNQ4 foi asociada inicialmente a unha forma de xordeira hereditaria. Curiosamente, un estudo recente localiza KCNQ4 nas terminacións nerviosas periféricas do folículo piloso de adaptación cutánea rápida e corpúsculo de Meissner. En consecuencia, a perda da función KCNQ4 leva a un aumento selectivo da sensibilidade dos mecaneceptoras á vibración de baixa frecuencia. En particular, as persoas con perda auditiva tardía debido a mutacións dominantes do xene KCNQ4 mostran un mellor rendemento na detección de vibracións de pequena amplitude e de pequena amplitude.
Insight do Dr. Alex Jimenez
O toque é considerado un dos sentidos máis complexos do corpo humano, especialmente porque non hai ningún organismo específico que o encargue. En lugar diso, o sentimento do tacto prodúcese a través de receptores sensoriais, coñecidos como mecanorreceptores, que se atopan en toda a pel e responden á presión ou distorsión mecánica. Hai catro tipos principais de mecanorreceptores na pel glabro ou sen pel dos mamíferos: corpúsculos lamelados, corpúsculos táctiles, terminacións nerviosas de Merkel e corpúsculos bulbosos. Os mecanorreceptores funcionan para permitir a detección do toque, para controlar a posición dos músculos, ósos e articulacións, coñecida como propiocepción, e mesmo para detectar sons e o movemento do corpo. Comprender os mecanismos de estrutura e función destes mecanorreceptores é un elemento fundamental na utilización de tratamentos e terapias para a xestión da dor.
Conclusión
O toque é un sentido complexo porque representa diferentes calidades táctiles, a vibración, a forma, a textura, o pracer e a dor, con distintas actuacións discriminatorias. Ata agora, a correspondencia entre un órgano táctil e o sentido psicofísico era correlativo e só aparecen marcadores moleculares de clase. O desenvolvemento de probas de roedores que coinciden coa diversidade de comportamentos táctiles son agora necesarios para facilitar a identificación futura da genómica. O uso de ratos que non posúen subconjuntos específicos de tipos aferentes sensoriais facilitará en gran medida a identificación de mecanorreceptores e fibras aferentes sensoriais asociadas a unha modalidade de toque particular. Curiosamente, un artigo recente abre a importante cuestión da base xenética dos trazos mecanosensuais en humanos e suxire que a mutación xenética única podería influír negativamente na sensibilidade no toque. 110 Isto subliña que a fisiopatoloxía do déficit de toque humano é en gran parte descoñecida e ciertamente descoñécese progreso identificando con precisión o subconxunto das neuronas sensoriais ligadas a unha modalidade táctil ou un déficit táctil.
En cambio, fixéronse progresos para definir as propiedades biofísicas das correntes mecano-gated.64 O desenvolvemento de novas técnicas nos últimos anos, permitindo o seguimento dos cambios de tensión nas membranas, mentres rexistra a corrente mecánica-gated, demostrou valioso método experimental para describir correntes mecanosensibles con adaptación rápida, intermedia e lenta (revisadas en Delmas e colaboradores) .66,111 O futuro será determinar o papel das propiedades actuais nos mecanismos de adaptación de mecanorreceptores funcionalmente diversos e da contribución das correntes mecanosensibles K + á excitabilidade de LTMRs e HTMRs.
A natureza molecular das correntes mecano-pechadas nos mamíferos é tamén un futuro tema de investigación prometedor. As investigacións futuras progresarán en dúas perspectivas, primeiro para determinar o papel da molécula accesoria que unen os canles ao citoesqueleto e requirirían para conferir ou regular a mecanosensibilidade das canles iónicas das familias TRP e ASIC / EnaC. En segundo lugar, investigar a área grande e prometedora da contribución dos canles piezoais, respondendo a cuestións clave, relativas aos mecanismos de permeación e peche, o subconxunto de neuronas sensoriais e modalidades táctiles que inclúen o piezo e o papel do piezo nas células non neuronais asociadas a mecanosensación.
O sentido do tacto, en comparación co da vista, o gusto, o son e o olfacto, que utilizan órganos específicos para procesar estas sensacións, pode ocorrer en todo o corpo a través de pequenos receptores coñecidos como mecanorreceptores. Pódense atopar diferentes tipos de mecanorreceptores en varias capas da pel, onde poden detectar unha gran variedade de estimulación mecánica. O artigo anterior describe aspectos destacados específicos que demostran o progreso dos mecanismos estruturais e funcionais dos mecanorreceptores asociados ao sentido do tacto. Información referenciada do Centro Nacional de Información Biotecnolóxica (NCBI). O alcance da nosa información limítase á quiropráctica, así como ás lesións e condicións da columna vertebral. Para discutir o tema, non dubide en preguntar ao Dr. Jiménez ou en contacto connosco en�915-850-0900 .
Comisariado polo Dr. Alex Jiménez
Temas adicionais: dor nas costas
Dor nas costas é unha das causas máis frecuentes de discapacidade e días perdidos no traballo en todo o mundo. En realidade, a dor lumbar atribúese como o segundo motivo máis frecuente para as consultas médicas, superadas en número por infeccións respiratorias superiores. Aproximadamente o 80 por cento da poboación experimentará algún tipo de dor nas costas polo menos unha vez ao longo da súa vida. A columna vertebral é unha estrutura complexa composta de ósos, articulacións, ligamentos e músculos, entre outros tecidos brandos. Por iso, feridas e / ou condicións agravadas, como discos herniados, pode levar a síntomas de dor nas costas. As lesións deportivas ou as lesións por accidentes automovilísticos adoitan ser a causa máis frecuente de dor nas costas, con todo, ás veces o movemento máis sinxelo pode ter resultados dolorosos. Afortunadamente, as opcións de tratamento alternativas, como o coidado quiropráctico, poden axudar a aliviar a dor nas costas mediante o uso de axustes espiñentos e manipulacións manuais, mellorando o alivio da dor.
1. Moriwaki K, Yuge O. Topographical features of cutaneous tactile hypoesthetic and hyperesthetic anomalies in chronic pain.Dor1999;81:1�6. doi: 10.1016/S0304-3959(98)00257-7.�[PubMed][Cruz Ref]
2. Shim B, Kim DW, Kim BH, Nam TS, Leem JW, Chung JM. Sensibilización mecánica e térmica de nociceptores cutáneos en ratas con neuropatía periférica experimental.Neurociencia...2005;132: 193-201. doi: 10.1016/j.neuroscience.2004.12.036.�[PubMed]�[Cruz Ref]
3. Kleggetveit IP, J�rum E. Disfunción das fibras grandes e pequenas nas lesións dos nervios periféricos con ou sen dor espontánea.J Dor2010;11: 1305�10. doi: 10.1016/j.jpain.2010.03.004.�[PubMed]�[Cruz Ref]
4. Noback CR. Morfoloxía e filoxenia do cabeloAnn NY Acad Sci1951;53:476�92. doi: 10.1111/j.1749-6632.1951.tb31950.x.�[PubMed]�[Cruz Ref]
5. Straile WE. Folículos pilosos protectores atípicos na pel do coelloNatureza1958;181:1604�5. doi: 10.1038/1811604a0.�[PubMed]�[Cruz Ref]
6. Straile WE. Morfoloxía dos folículos tilótricos na pel do coelloSon J Anat...1961;109: 1�13. doi: 10.1002/aja.1001090102.�[PubMed]�[Cruz Ref]
7. Millard CL, Woolf CJ. Inervación sensorial dos pelos da extremidade posterior da rata: unha análise microscópica ópticaJ Comp Neurol1988;277: 183-94. doi: 10.1002/cne.902770203.�[PubMed]�[Cruz Ref]
8. Hamann W. Mecanorreceptores cutáneos de mamíferos.�Prog Biophys Mol Biol.�1995;64:81�104. doi: 10.1016/0079-6107(95)00011-9.�[Review]�[PubMed]�[Cruz Ref]
9. Brown AG, Iggo A. Un estudo cuantitativo de receptores cutáneos e fibras aferentes no gato e coello.J Physiol.�1967;193: 707[Artigo gratuíto de PMC]�[PubMed]
10. Burgess PR, Petit D, Warren RM. Tipos de receptores na pel peluda dos gatos subministrados por fibras mielínicasJ Neurophysiol1968;31: 833[PubMed]
11. Driskell RR, Giangreco A, Jensen KB, Mulder KW, Watt FM. As células da papila dérmica Sox2-positivas especifican o tipo de folículo piloso na epiderme dos mamíferos.Desenvolvemento.�2009;136: 2815�23. doi: 10.1242/dev.038620.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
12. Hussein MA. O patrón xeral da disposición do folículo piloso na rata e no ratoJ Anat.1971;109: 307[Artigo gratuíto de PMC]�[PubMed]
13. Vielkind U, Hardy MH. Patróns cambiantes das moléculas de adhesión celular durante o desenvolvemento do folículo piloso do pelaxe do rato. 2. Morfoxénese do folículo nos mutantes capilares, Tabby e Downy.�Acta Anat (Basilea)�1996;157: 183-94. doi: 10.1159 / 000147880. [PubMed]�[Cruz Ref]
14. Hardy MH, Vielkind U. Patróns cambiantes de moléculas de adhesión celular durante o desenvolvemento do folículo piloso do pelaxe do rato. 1. Morfoxénese do folículo en ratos de tipo salvaxe.�Acta Anat (Basilea)�1996;157: 169-82. doi: 10.1159 / 000147879. [PubMed]�[Cruz Ref]
15. Li L, Rutlin M, Abraira VE, Cassidy C, Kus L, Gong S, et al. A organización funcional das neuronas mecanosensoriais cutáneas de baixo limiarCela2011;147: 1615�27. doi: 10.1016/j.cell.2011.11.027.[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
16. Brown AG, Iggo A. Un estudo cuantitativo de receptores cutáneos e fibras aferentes no gato e coello.J Physiol.�1967;193: 707[Artigo gratuíto de PMC]�[PubMed]
17. Burgess PR, Petit D, Warren RM. Tipos de receptores na pel peluda dos gatos subministrados por fibras mielínicasJ Neurophysiol1968;31: 833[PubMed]
18. Vallbo A, Olausson H, Wessberg J, Norrsell U. A system of unmyelinated afferents for innocuous mechanoreception in the human skin.Resumo cerebral1993;628:301�4. doi: 10.1016/0006-8993(93)90968-S.[PubMed]�[Cruz Ref]
19. Vallbo AB, Olausson H, Wessberg J. As aferentes non amielínicas constitúen un segundo sistema que codifica estímulos táctiles da pel peluda humana.J Neurophysiol1999;81: 2753[PubMed]
22. Wessberg J, Olausson H, Fernstr�m KW, Vallbo AB. Propiedades do campo receptivo das aferentes táctiles non mielínicas na pel humanaJ Neurophysiol2003;89: 1567-75. doi: 10.1152/jn.00256.2002.�[PubMed][Cruz Ref]
23. Liu Q, Vrontou S, Rice FL, Zylka MJ, Dong X, Anderson DJ. Visualización xenética molecular dun subconxunto raro de neuronas sensoriais non mielínicas que poden detectar un tacto suave.Nat Neurosci2007;10:946�8. doi: 10.1038/nn1937.�[PubMed]�[Cruz Ref]
24. Olausson H, Lamarre Y, Backlund H, Morin C, Wallin BG, Starck G, et al. As aferentes táctiles non mielínicas sinalan o tacto e proxectan ao córtex insularNat Neurosci2002;5: 900�4. doi: 10.1038/nn896.[PubMed]�[Cruz Ref]
25. Olausson H, Wessberg J, Morrison I, McGlone F, Vallbo A. The neurophysiology of unmyelinated tactile afferents.Neurosci Biobehav Rev. 2010;34: 185-91. doi: 10.1016/j.neubiorev.2008.09.011.�[Revisión][PubMed]�[Cruz Ref]
26. Kr�mer HH, Lundblad L, Birklein F, Linde M, Karlsson T, Elam M, et al. Activación da rede de dor cortical mediante estimulación táctil suave despois da inxección de sumatriptánDor2007;133: 72-8. doi: 10.1016 / j.pain.2007.03.001. [PubMed]�[Cruz Ref]
27. Applebaum AE, Beall JE, Foreman RD, Willis WD. Organización e campos receptivos das neuronas do tracto espinotalámico dos primatesJ Neurophysiol1975;38: 572[PubMed]
28. Branco JC, Sweet WH. Eficacia da cordotomía na dor pantasma despois da amputaciónAMA Arch Neurol Psiquiatría...1952;67: 315[PubMed]
29. Halata Z, Grim M, Bauman KI. Friedrich Sigmund Merkel e a súa "célula de Merkel", morfoloxía, desenvolvemento e fisioloxía: revisión e novos resultados.Anat Rec A Discov Mol Cell Evol Biol.�2003;271: 225. doi: 39/ar.a.10.1002.�[PubMed]�[Cruz Ref]
30. Morrison KM, Miesegaes GR, Lumpkin EA, Maricich SM. As células de Merkel de mamíferos descenden da liñaxe epidérmicaDev Biol.2009;336: 76. doi: 83/j.ydbio.10.1016.[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
31. Van Keymeulen A, Mascre G, Youseff KK, Harel I, Michaux C, De Geest N, et al. Os proxenitores epidérmicos orixinan células de Merkel durante o desenvolvemento embrionario e a homeostase do adulto.Bioloxía celular J2009;187: 91. doi: 100/jcb.10.1083.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
32. Ebara S, Kumamoto K, Baumann KI, Halata Z. As análises tridimensionais de cúpulas táctiles na pel peluda da pata de gato revelan substratos morfolóxicos para o procesamento sensorial complexo.Res. Neurociencia2008;61: 159. doi: 71/j.neures.10.1016.�[PubMed]�[Cruz Ref]
33. Guinard D, Usson Y, Guillermet C, Saxod R. Merkel complexos de pel dixital humana: imaxe tridimensional con microscopía láser confocal e inmunofluorescencia dobre.J Comp Neurol1998;398:98�104. doi: 10.1002/(SICI)1096-9861(19980817)398:1<98::AID-CNE6>3.0.CO;2-4.�[PubMed][Cruz Ref]
34. Reinisch CM, Tschachler E. A cúpula táctil na pel humana é subministrada por diferentes tipos de fibras nerviosas.Ann Neurol...2005;58: 88. doi: 95/ana.10.1002.�[PubMed]�[Cruz Ref]
35. Maricich SM, Morrison KM, Mathes EL, Brewer BM. Os roedores confían nas células de Merkel para tarefas de discriminación de texturasJ Neurosciencia2012;32:3296�300. doi: 10.1523/JNEUROSCI.5307-11.2012.[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
36. Ikeda I, Yamashita Y, Ono T, Ogawa H. A destrución fototóxica selectiva das células de Merkel de rata elimina as respostas de unidades de mecanorreceptores tipo I de adaptación lenta.J Physiol.�1994;479: 247[Artigo gratuíto de PMC][PubMed]
37. Maricich SM, Wellnitz SA, Nelson AM, Lesniak DR, Gerling GJ, Lumpkin EA, et al. As células de Merkel son esenciais para as respostas ao toque lixeiroCiencia2009;324: 1580�2. doi: 10.1126/science.1172890.[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
38. Diamond J, Holmes M, Enfermeira CA. As sinapses recíprocas entre células de Merkel e neuritas están implicadas no inicio das respostas táctiles na pel de salamandra?J Physiol.�1986;376: 101[Artigo gratuíto de PMC][PubMed]
39. Yamashita Y, Akaike N, Wakamori M, Ikeda I, Ogawa H. Correntes dependentes da tensión en células de Merkel illadas de ratas.J Physiol.�1992;450: 143[Artigo gratuíto de PMC]�[PubMed]
40. Wellnitz SA, Lesniak DR, Gerling GJ, Lumpkin EA. A regularidade do disparo sostido revela dúas poboacións de receptores táctiles de adaptación lenta na pel peluda do rato.J Neurophysiol2010;103: 3378-88. doi: 10.1152/jn.00810.2009.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
41. Nandasena BG, Suzuki A, Aita M, Kawano Y, Nozawa-Inoue K, Maeda T. Immunolocalization of aquaporin-1 in the mechanoreceptive Ruffini endings in the periodontal ligament.Resumo cerebral2007;1157: 32. doi: 40/j.brainres.10.1016.�[PubMed]�[Cruz Ref]
42. Rahman F, Harada F, Saito I, Suzuki A, Kawano Y, Izumi K, et al. Detección da canle iónica sensor de ácido 3 (ASIC3) en terminacións Ruffini periodontais dos incisivos do rato.Neurosci Lett. 2011;488: 173�7. doi: 10.1016/j.neulet.2010.11.023.�[PubMed]�[Cruz Ref]
43. Johnson KO. Papeis e funcións dos mecanorreceptores cutáneosCurr Opin Neurobiol2001;11:455�61. doi: 10.1016/S0959-4388(00)00234-8.�[Review]�[PubMed]�[Cruz Ref]
44. Wende H, Lechner SG, Cheret C, Bourane S, Kolanczyk ME, Pattyn A, et al. O factor de transcrición c-Maf controla o desenvolvemento e a función do receptor táctilCiencia2012;335: 1373�6. doi: 10.1126/science.1214314.�[PubMed]�[Cruz Ref]
45. Mendelson M, Lowenstein WR. Mecanismos de adaptación dos receptoresCiencia1964;144: 554�5. doi: 10.1126/science.144.3618.554.�[PubMed]�[Cruz Ref]
46. Loewenstein WR, Mendelson M. Components of receptor adaptation in a pacinian corpuscle.�J Physiol.�1965;177: 377[Artigo gratuíto de PMC]�[PubMed]
47. Pawson L, Prestia LT, Mahoney GK, G��l� B, Cox PJ, Pack AK. A interacción GABAérxico/glutamatérxico-glial/neuronal contribúe á rápida adaptación dos corpúsculos de Pacini.J Neurosciencia2009;29:2695�705. doi: 10.1523/JNEUROSCI.5974-08.2009.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
48. Basbaum AI, Jessell TM. A percepción da dor. En: Kandel ER, Schwartz JH, Jessell TM, eds. Principios da ciencia neuronal. Cuarta edición. Os compases de McGraw-Hill, 2000: 472-490.
49. Bourane S, Garces A, Venteo S, Pattyn A, Hubert T, Fichard A, et al. Os subtipos de mecanorreceptores de limiar baixo expresan selectivamente MafA e especifícanse mediante a sinalización Ret.Neuron2009;64: 857. doi: 70/j.neuron.10.1016.�[PubMed]�[Cruz Ref]
50. Kramer I, Sigrist M, de Nooij JC, Taniuchi I, Jessell TM, Arber S. A role for Runx transcription factor signaling in dorsal root ganglion sensory neuron diversification.Neuron2006;49: 379. doi: 93/j.neuron.10.1016.�[PubMed]�[Cruz Ref]
51. Luo W, Enomoto H, Rice FL, Milbrandt J, Ginty DD. Identificación molecular de mecanorreceptores de adaptación rápida e a súa dependencia do desenvolvemento da sinalización ret.�Neuron2009;64: 841. doi: 56/j.neuron.10.1016.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
52. Vallbo AB, Hagbarth KE. Actividade dos mecanorreceptores da pel rexistrada de forma percutánea en suxeitos humanos espertosExp Neurol.�1968;21:270�89. doi: 10.1016/0014-4886(68)90041-1.�[PubMed]�[Cruz Ref]
53. Macefield VG. Características fisiolóxicas dos mecanorreceptores de baixo limiar nas articulacións, músculos e pel en suxeitos humanos.Clin Exp Pharmacol Physiol.�2005;32:135�44. doi: 10.1111/j.1440-1681.2005.04143.x.�[Review]�[PubMed]�[Cruz Ref]
54. Koizumi S, Fujishita K, Inoue K, Shigemoto-Mogami Y, Tsuda M, Inoue K. As ondas de Ca2+ nos queratinocitos transmítense ás neuronas sensoriais: a implicación da activación do receptor ATP extracelular e P2Y2.Bioquímica J2004;380: 329. doi: 38/BJ10.1042.�[Artigo gratuíto de PMC]�[PubMed][Cruz Ref]
55. Azorin N, Raoux M, Rodat-Despoix L, Merrot T, Delmas P, Crest M. A sinalización de ATP é crucial para a resposta dos queratinocitos humanos á estimulación mecánica por choque hipoosmótico.Exp Dermatol.2011;20:401�7. doi: 10.1111/j.1600-0625.2010.01219.x.�[PubMed]�[Cruz Ref]
56. Amano M, Fukata Y, Kaibuchi K. Regulación e funcións da quinase Rho-associated.Exp Cell Res.�2000;261: 44. doi: 51/excr.10.1006.�[Revisión]�[PubMed]�[Cruz Ref]
57. Koyama T, Oike M, Ito Y. Implicación da Rho-quinase e a tirosina quinase na liberación de ATP inducida por estrés hipotónico en células endoteliais aórticas bovinas.J Physiol.�2001;532:759�69. doi: 10.1111/j.1469-7793.2001.0759e.x.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
59. McCarter GC, Reichling DB, Levine JD. Transdución mecánica por neuronas ganglionares da raíz dorsal de rata in vitroNeurosci Lett. 1999;273:179�82. doi: 10.1016/S0304-3940(99)00665-5.�[PubMed]�[Cruz Ref]
60. Drew LJ, Wood JN, Cesare P. Distinct mechanosensitive properties of capsaicin-sensitive and -insensitive sensory neurons.J Neurosciencia2002;22:RC228.�[PubMed]
61. Drew LJ, Rohrer DK, Price MP, Blaver KE, Cockayne DA, Cesare P, et al. As canles iónicas sensoriais de ácido ASIC2 e ASIC3 non contribúen ás correntes activadas mecánicamente nas neuronas sensoriais dos mamíferos.J Physiol.�2004;556: 691-710. doi: 10.1113/jphysiol.2003.058693.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
62. McCarter GC, Levine JD. Base iónica dunha corrente de mecanotransdución en neuronas ganglionares da raíz dorsal de rata adulta.Mol Pain2006;2:28. doi: 10.1186/1744-8069-2-28.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
63. Coste B, Crest M, Delmas P. Disección farmacolóxica e distribución de correntes de NaN/Nav1.9, Ca2+ tipo T e correntes catiónicas activadas mecánicamente en diferentes poboacións de neuronas DRG.�J Gen Physiol.�2007;129: 57. doi: 77/jgp.10.1085.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
64. Hao J, Delmas P. Os mecanismos de desensibilización múltiple das canles mecanotransdutores dan forma ao disparo das neuronas mecanosensoriais.J Neurosciencia2010;30:13384�95. doi: 10.1523/JNEUROSCI.2926-10.2010.[PubMed]�[Cruz Ref]
65. Drew LJ, Wood JN. FM1-43 é un bloqueador permanente das canles iónicas mecanosensibles nas neuronas sensoriais e inhibe as respostas do comportamento aos estímulos mecánicos.Mol Pain2007;3:1. doi: 10.1186/1744-8069-3-1.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
66. Hao J, Delmas P. Gravación de correntes mecanosensibles mediante un mecanoestimulador accionado por piezoeléctricos.Nat Protoc.�2011;6: 979-90. doi: 10.1038/nprot.2011.343.�[PubMed]�[Cruz Ref]
67. Rugiero F, Drew LJ, Wood JN. Propiedades cinéticas das correntes activadas mecánicamente nas neuronas sensoriais da columna vertebralJ Physiol.�2010;588: 301-14. doi: 10.1113/jphysiol.2009.182360.�[Artigo gratuíto de PMC]�[PubMed][Cruz Ref]
68. Hu J, Lewin GR. Correntes mecanosensibles nas neuritas de neuronas sensoriais de rato cultivadasJ Physiol.�2006;577: 815-28. doi: 10.1113/jphysiol.2006.117648.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
69. Bhattacharya MR, Bautista DM, Wu K, Haeberle H, Lumpkin EA, Julius D. O estiramento radial revela poboacións distintas de neuronas somatosensoriales de mamíferos mecanosensibles.Proc Natl Acad Sci US A. 2008;105: 20015-20. doi: 10.1073/pnas.0810801105.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
70. Crawford AC, Evans MG, Fettiplace R. Activation and adaptation of transducer currents in turtle hair cells.J Physiol.�1989;419: 405[Artigo gratuíto de PMC]�[PubMed]
71. Ricci AJ, Wu YC, Fettiplace R. The endogenous calcium buffer and the time course of transducer adaptation in auditive hair cells.J Neurosciencia1998;18: 8261[PubMed]
72. Vollrath MA, Kwan KY, Corey DP. A micromaquinaria de mecanotransdución nas células ciliadasAnnu Rev Neurosci2007;30: 339. doi: 65/annurev.neuro.10.1146.�[Revisión]�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
73. Goodman MB, Schwarz EM. Transdución do tacto en Caenorhabditis elegansAnnu Rev Physiol.�2003;65: 429. doi: 52/annurev.physiol.10.1146.�[Revisión]�[PubMed]�[Cruz Ref]
74. Waldmann R, Lazdunski MH. Canles de cationes H(+): sensores de ácidos neuronais na familia de canles iónicos NaC/DEG.Curr Opin Neurobiol1998;8:418�24. doi: 10.1016/S0959-4388(98)80070-6.[Review]�[PubMed]�[Cruz Ref]
75. Page AJ, Brierley SM, Martin CM, Martinez-Salgado C, Wemmie JA, Brennan TJ, et al. A canle iónica ASIC1 contribúe á función mecanorreceptora visceral pero non cutáneaGastroenteroloxía2004;127: 1739-47. doi: 10.1053/j.gastro.2004.08.061.�[PubMed]�[Cruz Ref]
76. Price MP, McIlwrath SL, Xie J, Cheng C, Qiao J, Tarr DE, et al. A canle catiónica DRASIC contribúe á detección do tacto cutáneo e dos estímulos ácidos en ratos.Neuron2001;32: 1071-83. doi: 10.1016/S0896-6273(01)00547-5.�[Erratum en: Neuron 2002 Jul 18;35] [2]�[PubMed]�[Cruz Ref]
77. Roza C, Puel JL, Kress M, Baron A, Diochot S, Lazdunski M, et al. A eliminación da canle ASIC2 en ratos non prexudica a mecanosensación cutánea, a mecanocicepción visceral e a audición.J Physiol.�2004;558: 659-69. doi: 10.1113/jphysiol.2004.066001.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
78. Damann N, Voets T, Nilius B. TRPs in our senses.�Curr Biol.�2008;18: R880�9. doi: 10.1016/j.cub.2008.07.063.�[Revisión]�[PubMed]�[Cruz Ref]
79. Christensen AP, Corey DP. Canles TRP na mecanosensación: ¿activación directa ou indirecta?Nat Rev Neurosci2007;8: 510�21. doi: 10.1038/nrn2149.�[Revisión]�[PubMed]�[Cruz Ref]
80. Liedtke W, Tobin DM, Bargmann CI, Friedman JM. TRPV4 de mamíferos (VR-OAC) dirixe as respostas comportamentais a estímulos osmóticos e mecánicos en Caenorhabditis elegans.Proc Natl Acad Sci US A. 2003;100(Suplemento 2): 14531�6. doi: 10.1073/pnas.2235619100.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
81. Suzuki M, Mizuno A, Kodaira K, Imai M. Sensación de presión deteriorada en ratos que carecen de TRPV4.J Biol Chem2003;278: 22664�8. doi: 10.1074/jbc.M302561200.�[PubMed]�[Cruz Ref]
82. Liedtke W, Choe Y, Mart�-Renom MA, Bell AM, Denis CS, Sali A, et al. Canle activado osmóticamente relacionado co receptor vaniloide (VR-OAC), un candidato a osmoreceptor de vertebrados.Cela2000;103:525�35. doi: 10.1016/S0092-8674(00)00143-4.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
83. Alessandri-Haber N, Dina OA, Yeh JJ, Parada CA, Reichling DB, Levine JD. O potencial receptor transitorio do vaniloide 4 é esencial na dor neuropática inducida pola quimioterapia na rataJ Neurosciencia2004;24: 4444-52. doi: 10.1523/JNEUROSCI.0242-04.2004.�[Erratum en: J Neurosci. xuño de 2004; 24] [23][PubMed]�[Cruz Ref]
84. Bautista DM, Jordt SE, Nikai T, Tsuruda PR, Read AJ, Poblete J, et al. TRPA1 media as accións inflamatorias de irritantes ambientais e axentes proalxésicosCela2006;124: 1269-82. doi: 10.1016/j.cell.2006.02.023.�[PubMed]�[Cruz Ref]
85. Kwan KY, Allchorne AJ, Vollrath MA, Christensen AP, Zhang DS, Woolf CJ, et al. TRPA1 contribúe á nocicepción fría, mecánica e química, pero non é esencial para a transdución das células ciliadas.Neuron2006;50: 277. doi: 89/j.neuron.10.1016.�[PubMed]�[Cruz Ref]
86. Coste B, Mathur J, Schmidt M, Earley TJ, Ranade S, Petrus MJ, et al. Piezo1 e Piezo2 son compoñentes esenciais de distintas canles catiónicas activadas mecánicamenteCiencia2010;330: 55�60. doi: 10.1126/science.1193270.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
87. Coste B, Xiao B, Santos JS, Syeda R, Grandl J, Spencer KS, et al. As proteínas piezoeléctricas son subunidades formadoras de poros de canles activadas mecánicamenteNatureza2012;483: 176. doi: 81/nature10.1038.[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
88. Bae C, Sachs F, Gottlieb PA. A canle iónica mecanosensible Piezo1 está inhibida polo péptido GsMTx4.�Bioquímica...2011;50: 6295-300. doi: 10.1021/bi200770q.�[Artigo gratuíto de PMC]�[PubMed][Cruz Ref]
89. Kim SE, Coste B, Chadha A, Cook B, Patapoutian A. The role of Drosophila Piezo in mechanical nociception.Natureza2012;483: 209�12. doi: 10.1038/nature10801.�[Artigo gratuíto de PMC]�[PubMed][Cruz Ref]
90. Zarychanski R, Schulz VP, Houston BL, Maksimova Y, Houston DS, Smith B, et al. As mutacións na proteína de mecanotransdución PIEZO1 están asociadas á xerocitose hereditaria.Sangue.2012;120:1908�15. doi: 10.1182/blood-2012-04-422253.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
91. Kawashima Y, G�l�oc GS, Kurima K, Labay V, Lelli A, Asai Y, et al. A mecanotransdución nas células ciliadas do oído interno do rato require xenes parecidos a canles transmembranaJ Clin Invest2011;121: 4796-809. doi: 10.1172/JCI60405.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
92. Tlili A, Rebeh IB, Aifa-Hmani M, Dhouib H, Moalla J, Tlili-Chouch�ne J, et al. TMC1 pero non TMC2 é responsable da deficiencia auditiva autosómica recesiva non sindrómica nas familias tunisianas.Audiol Neurootol...2008;13: 213-8. doi: 10.1159 / 000115430. [PubMed]�[Cruz Ref]
93. Manji SS, Miller KA, Williams LH, Dahl HH. Identificación de tres novas cepas de rato con hipoacusia con mutacións no xene Tmc1Son J Pathol...2012;180: 1560�9. doi: 10.1016/j.ajpath.2011.12.034.[PubMed]�[Cruz Ref]
94. Wetzel C, Hu J, Riethmacher D, Benckendorff A, Harder L, Eilers A, et al. Unha proteína do dominio de estomatina esencial para a sensación táctil no ratoNatureza2007;445: 206�9. doi: 10.1038/nature05394.�[PubMed][Cruz Ref]
95. Martínez-Salgado C, Benckendorff AG, Chiang LY, Wang R, Milenkovic N, Wetzel C, et al. Mecanotransdución da estomatina e das neuronas sensoriaisJ Neurophysiol2007;98: 3802�8. doi: 10.1152/jn.00860.2007.[PubMed]�[Cruz Ref]
96. Huang M, Gu G, Ferguson EL, Chalfie M. Unha proteína similar á estomatina necesaria para a mecanosensación en C. elegans.Natureza1995;378:292�5. doi: 10.1038/378292a0.�[PubMed]�[Cruz Ref]
97. Hu J, Chiang LY, Koch M, Lewin GR. Evidencia dunha atadura proteica implicada no tacto somáticoEMBO J.�2010;29: 855-67. doi: 10.1038/emboj.2009.398.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
98. Chiang LY, Poole K, Oliveira BE, Duarte N, Sierra YA, Bruckner-Tuderman L, et al. A laminina-332 coordina a mecanotransdución e a bifurcación do cono de crecemento nas neuronas sensoriais.Nat Neurosci2011;14: 993-1000. doi: 10.1038/nn.2873.�[PubMed]�[Cruz Ref]
99. Lesage F, Guillemare E, Fink M, Duprat F, Lazdunski M, Romey G, et al. TWIK-1, un ser humano omnipresente que se rectifica de forma débil cara a dentro dunha canle de K+ cunha estrutura nova.EMBO J.�1996;15: 1004 11.[Artigo gratuíto de PMC]�[PubMed]
100. Lesage F. Farmacoloxía das canles de potasio de fondo neuronal.�Neurofarmacoloxía...2003;44:1�7. doi: 10.1016/S0028-3908(02)00339-8.�[Review]�[PubMed]�[Cruz Ref]
101. Medhurst AD, Rennie G, Chapman CG, Meadows H, Duckworth MD, Kelsell RE, et al. Análise da distribución dos canles de potasio do dominio de dous poros humanos nos tecidos do sistema nervioso central e da periferia.Brain Res Mol Brain Res.�2001;86:101�14. doi: 10.1016/S0169-328X(00)00263-1.�[PubMed][Cruz Ref]
102. Maingret F, Patel AJ, Lesage F, Lazdunski M, Honor� E. Mechano- or acid stimulation, two interactive modes of activation of the TREK-1 potassium channel.�J Biol Chem1999;274: 26691�6. doi: 10.1074/jbc.274.38.26691.�[PubMed]�[Cruz Ref]
103. Maingret F, Fosset M, Lesage F, Lazdunski M, Honor� E. TRAAK é unha canle de K+ neuronal de mamíferos mechano-gate.�J Biol Chem1999;274: 1381�7. doi: 10.1074/jbc.274.3.1381.�[PubMed][Cruz Ref]
104. Alloui A, Zimmermann K, Mamet J, Duprat F, No�l J, Chemin J, et al. TREK-1, unha canle de K+ implicada na percepción polimodal da dorEMBO J.�2006;25: 2368-76. doi: 10.1038/sj.emboj.7601116.[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
105. No�l J, Zimmermann K, Busserolles J, Deval E, Alloui A, Diochot S, et al. As canles K+ activadas por mecanos TRAAK e TREK-1 controlan tanto a percepción cálida como fría.EMBO J.�2009;28: 1308-18. doi: 10.1038/emboj.2009.57.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
106. Dobler T, Springauf A, Tovornik S, Weber M, Schmitt A, Sedlmeier R, et al. As canles K+ de dous poros de TRESK constitúen un compoñente significativo das correntes de potasio de fondo nas neuronas do ganglio da raíz dorsal murina.J Physiol.�2007;585: 867-79. doi: 10.1113/jphysiol.2007.145649.[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
107. Bautista DM, Sigal YM, Milstein AD, Garrison JL, Zorn JA, Tsuruda PR, et al. Os axentes picantes dos pementos de Szechuan excitan as neuronas sensoriais ao inhibir as canles de potasio de dous poros.Nat Neurosci2008;11: 772-9. doi: 10.1038/nn.2143.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
108. Lennertz RC, Tsunozaki M, Bautista DM, Stucky CL. Bases fisiolóxicas da parestesia con formigueo evocada por hidroxi-alfa-sanshool.J Neurosciencia2010;30:4353�61. doi: 10.1523/JNEUROSCI.4666-09.2010.�[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
109. Heidenreich M, Lechner SG, Vardanyan V, Wetzel C, Cremers CW, De Leenheer EM, et al. As canles KCNQ4 K(+) sintonizan os mecanorreceptores para unha sensación táctil normal no rato e no home.Nat Neurosci2012;15: 138-45. doi: 10.1038/nn.2985.�[PubMed]�[Cruz Ref]
110. Frenzel H, Bohlender J, Pinsker K, Wohlleben B, Tank J, Lechner SG, et al. Unha base xenética para os trazos mecanosensoriais en humanosPLoS Biol.�2012;10: e1001318. doi: 10.1371 / journal.pbio.1001318.[Artigo gratuíto de PMC]�[PubMed]�[Cruz Ref]
111. Delmas P, Hao J, Rodat-Despoix L. Molecular mechanisms of mechanotransduction in mammalian sensory neurons.Nat Rev Neurosci2011;12: 139. doi: 53/nrn10.1038.�[PubMed]�[Cruz Ref]
Dor mecanorreceptiva: Segundo os CDC, máis do 50% dos adultos estadounidenses (125 millóns) tiñan unha enfermidade musculoesquelética. trastorno da dor en 2012.�
�Máis do 40% dos adultos con dor musculoesquelética utilizaron un enfoque complementario de saúde por calquera motivo en 2012. Isto foi significativamente maior que o uso entre persoas sen dor musculoesquelética (24.1%). O uso de enfoques complementarios de saúde por calquera motivo entre persoas con dor ou problemas de pescozo foi máis do dobre que o uso entre persoas sen estes problemas.
"Entre os adultos con trastorno de dor musculoesquelética, o uso de calquera enfoque complementario de saúde foi maior entre aqueles con dor ou problemas de pescozo (50.6%), seguidos por persoas con outros problemas musculoesqueléticos (46.2%).
�O uso de enfoques complementarios de saúde por calquera motivo entre persoas con dor ou problemas de pescozo foi máis do dobre que o uso entre persoas sen estes problemas.�
Os mecanorreceptores son receptores sensoriais que responden á presión mecánica ou á distorsión.
Estes inclúen receptores cutáneos para o tacto, receptores que controlan a lonxitude e tensión muscular, receptores auditivos e vestibulares e outros.
TEORÍA DO CONTROL DA PORTA DA DOR
A entrada non dolorosa pecha as portas á entrada dolorosa.
Isto evita que as sensacións de dor viaxan a niveis corticais máis altos
As aferentes de pequeno diámetro (dor) impiden a inhibición da dor
As aferentes de gran diámetro (vibración) tenden a excitar a inhibición da dor.
Esta teoría afirma que as fibras non nociceptivas poden interferir cos sinais das fibras da dor, polo tanto, inhibindo a dor.
Gran diámetro A? as fibras non son nociceptivas (non transmiten estímulos de dor) e inhiben os efectos do disparo por A? e fibras C.
COLUMNA DORSAL VÍA LEMNISCAL MEDIAL
UTILIZACIÓN DE MECANORECEPTORES PERIFÉRICOS PARA ALTERAR A PERCEPCIÓN DA DOR
COMO PODEMOS AXUDAR?
REFLEXO DE RETIRO
Unha neurona aferente excitada estimula as interneuronas excitatorias que á súa vez estimulan as neuronas motoras eferentes que irrigan o bíceps, o músculo do brazo que flexiona (dobra) a articulación do cóbado. A contracción do bíceps afasta a man da cociña quente.
A neurona aferente tamén estimula as interneuronas inhibidoras que á súa vez inhiben as neuronas eferentes que irrigan o tríceps para evitar que se contraiga. Este tipo de conexión neuronal que implica a estimulación da subministración nerviosa a un músculo e a inhibición simultánea dos nervios ao seu músculo antagónico coñécese como inhibición recíproca.
A neurona aferente aínda estimula outras interneuronas que transportan o sinal ata o cerebro pola medula espiñal a través dunha vía ascendente. Só cando o impulso chega á zona sensorial do córtex é consciente da dor, da súa localización e do tipo de estímulo. Ademais, cando o impulso chega ao cerebro, a información pódese almacenar como memoria e a persoa pode pensar no que pasou.
TERAPIA BASADA EN RECEPTORES
Axustes
A activación dos mecanorreceptores articulares mediante axustes quiroprácticos pode modular e "sombrear" a percepción cerebral de fibras de menor diámetro.
A repetición da activación dos mecanorreceptores articulares pode crear plasticidade positiva nas vías aferentes.
A plasticidade positiva pode apagar a dor
Vibración
A estimulación vibratoria a frecuencias específicas pode alterar a percepción da dor
A repetición da activación dos discos de Merkel e dos corpúsculos de Meissner pode crear plasticidade positiva nas vías aferentes.
De novo, a plasticidade positiva pode apagar a dor
Vibrado
�Este tipo de dispositivo aplica vibracións sinusoidais e ofrece unha amplitude seleccionable continua de 0-5.2 mm dependendo da posición do pé e unha frecuencia seleccionable de 5-30 Hz.�
�O adestramento de WBV parece ser unha intervención eficaz, segura e adecuada para os empregados que traballan sentados con dor lumbar crónica.
A estimulación vibrotáctil homotópica resultou nunha redución da dor térmica do 40% en todos os grupos de temas. A distracción non pareceu afectar as valoracións experimentais da dor
�A estimulación vibrotáctil recrutou de forma eficaz mecanismos analxésicos non só en NC senón tamén en pacientes con dor musculoesquelética crónica, incluíndo FM.�
TOCO DE LUZ
En total, 44 voluntarios sans experimentaron dor térmica e estímulos óptimos de TC (cepillado lento) e subóptimos de TC (cepillado rápido ou vibración). Utilizáronse tres paradigmas experimentais diferentes: Aplicación concomitante de dor térmica e estimulación táctil (cepillado lento ou vibración); Cepillado lento, aplicado por duración e intervalos variables, que precede a dor de calor; Cepillado lento versus rápido que precede a dor de calor
Nos humanos, as principais áreas cerebrais que reciben información C-LTMR pertencen ao sistema somatosensorial e afectan ás redes cerebrais de procesamento como a cortiza insular posterior contralateral ou a cortiza prefrontal medial. A intensidade do tacto dirixido á TC está codificada no córtex somatosensorial primario e secundario (S1 contralateral, S2 bilateral), mentres que a agradableidade está codificada no córtex cingulado anterior. Os C-LTMR tamén activan rexións implicadas no procesamento da recompensa (putamen e cortiza orbitofrontal) e no procesamento de estímulos sociais (sulco temporal superior posterior).
TODO O PERIFÉRICO TEN UNHA CONSECUENCIA CENTRAL
ESTUDO DE CASO
Un home de 47 anos sufriu CVA esquerda en outubro de 2017.
Non moveu o lado dereito do seu corpo desde o accidente.
Presentouse na nosa clínica porque quere "volver a ela".
DESTACADOS DE EXAMENO FÍSICO
Disartria
Percepción alterada da dor
Dificultade con matemáticas sinxelas
Flácido en RUE e RLE
DESTACADOS DE EXAMENO FÍSICO
O paciente non tiña movemento ata que comezamos a probar a sensación e os reflexos:
ALODINIA:�Refírese á sensibilización central da dor (aumento da resposta das neuronas) despois dunha estimulación normalmente non dolorosa e moitas veces repetitiva.
A alodinia pode provocar o desencadeamento dunha resposta de dor a partir de estímulos, que normalmente non provoca dor.
A temperatura ou os estímulos físicos poden provocar alodinia, que pode sentirse como unha sensación de ardor e adoita ocorrer despois da lesión nun sitio.
A aldinia é diferente da hiperalxesia, unha reacción extrema e esaxerada a un estímulo, que normalmente é dolorosa.
INTERVENCIÓNS TERAPÉUTICAS
Vibración
Toque Lixeiro
Acupressure
Frecuencias acústicas
Axustes!
DESPOIS DE DOUS DÍAS
Terapia baseada en receptores e dor mecanorreceptiva
Por que o dano ou lesión localizado causado polo trauma levan a dor crónica e intratável en determinados pacientes? ¿Que é o encargado da tradución de lesións locais con dor aguda nunha enfermidade crónica? ¿Por que a dor responde a medicamentos e / ou medicamentos antiinflamatorios, mentres que outras formas de dor requiren opiáceos?
Dor é un proceso intrincado que implica tanto o sistema nervioso periférico (PNS) como o sistema nervioso central (CNS). A lesión por tecido dispara o PNS, que transmite sinais a través da medula espiñal no cerebro, onde se produce a percepción da dor. Non obstante, o que fai que a intensa experiencia da dor se converta nun fenómeno incesante? Pódese facer algo para evitar isto? A evidencia indica iso Dor crónica resulta dunha combinación de mecanismos, como "recordos" neurolóxicos da dor anterior.
Nociception: The Simplest Pathway
A dor aguda ou nociceptiva caracterízase pola experiencia habitual de incomodidade que ocorre en resposta a danos ou lesións moi básicas. É protector, avisándonos para afastarnos da orixe do insulto e coidar do trauma. Os mecanismos que crean dor nociceptiva inclúen a transducción, que amplía a estimulación traumática externa á actividade eléctrica en nervios aferentes primarios especializados nociceptivos. Os nervios aferentes logo levan a información sensorial do SNP ao CNS.
No SNC, os datos de dor transmítense nas neuronas sensoriais primarias en células de proxección central. Despois de que a información se transfire a todas as áreas do cerebro que son responsables da nosa percepción, a experiencia sensorial real ocorre. A dor nociceptiva é unha reacción relativamente sinxela a un estímulo particularmente sinxelo e agudo. Pero os mecánicos encargados da dor nociceptiva non poden identificar fenómenos, como a dor que persiste a pesar da eliminación ou curación da estimulación, como por exemplo a dor de extremidades fantasmagóricas.
A dor ea resposta inflamatoria
En circunstancias de lesións máis graves, como as feridas cirúrxicas, o dano tisular pode estimular unha reacción inflamatoria. Non obstante, outras condicións, especialmente a artrite, tamén poden ser caracterizadas por casos continuos de inflamación asociados a síntomas intensos de dor. Os mecanismos para este tipo de dor relacionados co dano tisular e unha resposta inflamatoria son diferentes da dor nociceptiva de alerta temprana.
Observando a incisión ou o sitio de outro dano ou lesión, ocorre unha fervenza de eventos hiperexcitábeis no sistema nervioso. Este fenómeno "ventral" corporal comeza na pel, onde se potencia ao longo dos nervios periféricos e culmina cunha resposta de hipersensibilidad ao longo da médula espinal (dorsal) e ao cerebro. As células inflamatorias circundan as rexións do dano tisular e tamén producen citocinas e quimiocinas, substancias que pretenden mediar o proceso de curación e rexeneración do tecido. Pero estes axentes tamén poden considerarse irritantes e axustar as propiedades das neuronas sensoriais primarias en torno á área de trauma.
Así, os principais factores que desencadean a dor inflamatoria inclúen danos nos nociceptores de alto limiar, coñecidos como sensibilización periférica, cambios e alteracións das neuronas no sistema nervioso e a amplificación da excitabilidade das neuronas no SNC. Isto representa a sensibilización central e é responsábel pola hipersensibilidade, onde as áreas adxacentes ás da lesión real experimentarán dor coma se estivesen feridas. Estes tecidos tamén poden reaccionar á estimulación que normalmente non crea dor, como un toque, vestindo roupa, presión lixeira, ou mesmo cepillando o seu propio cabelo, coma se fosen realmente dolorosos, referidos como alodinia.
A dor neuropática resulta dun dano ou ferida ao sistema nervioso, como a síndrome do túnel do carpo, a neurálxica postéptica ea neuropatia diabética. Aínda que algúns dos mecanismos que parecen provocar a dor neuropática se solapan cos responsables da dor inflamatoria, moitos deles son diferentes e, polo tanto, necesitarán un enfoque diferente para a súa xestión.
O proceso de sensibilización periférica e central mantense, polo menos teoricamente e experimentalmente, durante o neurotransmisor excitatorio, glutamato, que se cre que se libera cando o receptor N-metil-D-aspartato (NMDA) está activado.
O sistema nervioso está composto por neurotransmisores inhibitorios ou excitatorios. A maior parte do que permite que o noso sistema nervioso responda de forma adecuada aos danos ou lesións é a afinación ou a inhibición dunha variedade de procesos. A overexcitación do sistema nervioso é considerada un problema en diferentes trastornos. Por exemplo, a sobreactivación dun receptor NMDA tamén pode estar relacionada con trastornos afectos, anormalidades simpáticas e mesmo a tolerancia ao opiáceo.
Incluso a dor nociceptiva ordinaria, ata certo punto, activa o receptor NMDA e crese que conduce á liberación de glutamato. Non obstante, na dor neuropática, a hipersensibilidade ao receptor NMDA é fundamental.
Con outros tipos de dor crónica, como a fibromialxia e dores de cabeza tipo tensión, algúns dos mecanismos activos na dor inflamatoria e neuropática tamén poden crear anomalías similares no sistema de dor, incluíndo a sensibilización central, maior excitabilidade das vías somatosensoriales e reducións mecanismos inhibidores do sistema nervioso central.
Sensibilización periférica
A ciclooxigenasa (COX) tamén desempeña unha función esencial nas sensibilizacións tanto periféricas como centrais. COX-2 é unha das enzimas que se induce durante o proceso inflamatorio; O COX-2 converte o ácido araquidónico en prostaglandinas, o que aumenta a sensibilidade dos terminales nociceptor periféricos. Practicamente, a inflamación periférica tamén causa que o COX-2 sexa producido a partir do SNC. As sinais dos nociceptores periféricos son parcialmente responsables deste upregulation, pero tamén parece haber un compoñente humoral para a transducción dos sinais de dor na barreira hematoencefálica.
Por exemplo, en modelos experimentais, a COX-2 xérase a partir do CNS mesmo se os animais reciben un bloqueo nervioso sensorial antes da estimulación inflamatoria periférica. O COX-2 que se expresa sobre as neuronas do corno dorsal da medula espiñal libera prostaglandinas, que actúan nos terminales centrais ou nas terminacións presinápticas das fibras sensoriais nociceptivas para aumentar a liberación do transmisor. Adicionalmente, actúan postsináticamente nas neuronas do corno dorsal para producir despolarización directa. E finalmente, inhiben a actividade do receptor de glicina, e este é un transmisor inhibitorio. Polo tanto, as prostaglandinas crean un aumento na excitabilidade das neuronas centrais.
Plasticidade cerebral e sensibilización central
A sensibilización central describe os cambios que ocorren no cerebro en reacción á estimulación nerviosa repetida. Tras repetir estímulos, as cantidades de hormonas e signos eléctricos cerebrais cambian a medida que as neuronas desenvolven unha "memoria" para reaccionar a estes signos. A estimulación constante crea unha memoria cerebral máis poderosa, polo que o cerebro responderá de forma máis rápida e efectiva ao pasar por unha estimulación idéntica no futuro. As modificacións consecuentes na fiación cerebral e reacción son coñecidas como plasticidade neuronal, que describen a capacidade do cerebro para alterarse facilmente ou a sensibilización central. Polo tanto, o cerebro é activado ou sensibilizado por estímulos previos ou repetidos para facerse máis excitável.
As fluctuacións da sensibilización central prodúcense despois de repetidas encontros con dor. A investigación nos animais indica que a exposición repetida a unha estimulación dolorosa cambiará o limiar da dor do animal e levará a unha resposta máis forte á dor. Os investigadores pensan que estas modificacións poden explicar a dor persistente que pode ocorrer mesmo despois da cirurxía traseira exitosa. Aínda que se pode eliminar un disco herniador dun nervio axustado, a dor pode continuar como unha memoria da compresión nerviosa. Os recentemente nados sometidos a circuncisión sen anestesia reaccionarán máis profundamente á futura estimulación dolorosa, como inxeccións de rutina, vacinas e outros procesos dolorosos. Estes nenos non só teñen unha reacción hemodinámica máis elevada, coñecida como taquicardia e taquipnea, pero tamén desenvolverán un arrepio maior.
Esta memoria neurológica da dor foi estudada extensamente. Nun informe sobre os seus estudos de investigación anteriores, Woolf observou que a excitabilidade reflexiva mellorada despois do dano ou lesión nos tecidos periféricos non depende de sinais de entrada periféricos continuados; máis ben, horas despois dun trauma periférico, os campos receptivos da neurona espinal dorsal continuaron aumentando. Os investigadores tamén documentaron o significado do receptor da NMDA espinal na inducción e mantemento da sensibilización central.
Importancia para a xestión da dor
Unha vez que se establece a sensibilización central, moitas veces son necesarios doses máis grandes de analxésicos para reprimirlo. A analgesia preventiva ou a terapia antes da progresión da dor poden reducir os efectos de todas estas estimulacións no SNC. Woolf demostrou que a dose de morfina necesaria para deter a hiperexcitabilidade central, dada antes dunha curta estimulación eléctrica nociva nas ratas, era unha décima dose necesaria para abolir a actividade despois de crecer. Isto tradúcese na práctica clínica.
Nun estudo clínico de pacientes con 60 sometidos a histerectomia abdominal, os individuos que recibiron 10 mg de morfina por vía intravenosa no momento da indución da anestesia requiriron unha morfina significativamente menor para o control post-operatorio da dor. Ademais, a sensibilidade á dor da ferida, referida como hiperalgesia secundaria, tamén se reduciu no grupo pretratado por morfina. A analxésia preventiva utilizouse cun éxito comparable nunha variedade de opcións cirúrxicas, incluíndo operacións prespartidas e operación post-ortopédica.
Unha soa dose de 40 ou 60 mg / kg de acetaminofeno rectal ten un claro efecto de limpeza da morfina na cirurxía de día en nenos, se se administra na inducción da anestesia. Ademais, os nenos con analgesia suficiente con acetaminofeno experimentaron náuseas e vómitos significativamente menos postoperatorios.
Os antagonistas dos receptores de NMDA impartiron analgesia postoperatoria cando se administran preoperatoriamente. Existen varios informes na literatura que apoian o uso de ketamina e dextrometorfano no período preoperatorio. Nos pacientes sometidos a reconstrución do ligamento cruzado anterior, o consumo de opioides por analoxia controlada por pacientes con 24-hora foi significativamente menor na categoría de dextrometorfano preoperatorio versus o grupo placebo.
En estudos de investigación dobre-cego e controlado con placebo, gabapentina foi indicado como analgésico premedicante para pacientes sometidos a mastectomía e histerectomía. A gabapentina oral preoperatoria reduciu as puntuacións da dor e o consumo analxésico postoperatorio sen separación dos efectos secundarios en comparación co placebo.
A administración preoperatoria de fármacos antiinflamatorios non esteroides (AINE) demostrou unha diminución significativa no uso opioide postoperatorio. Os COX-2 son preferibles debido á súa relativa falta de efectos plaquetarios e de perfil de seguridade gastrointestinal significativo cando se compara cos AINE convencionais. Celecoxib, rofecoxib, valdecoxib e parecoxib, fóra dos Estados Unidos, administraron preoperatoriamente o uso de narcóticos postoperatorios por máis de 40 por cento, con moitos pacientes que usan menos da metade dos opiáceos en comparación co placebo.
O bloqueo da condución nerviosa no período preoperatorio parece impedir o desenvolvemento da sensibilización central. Atribuíuse a síndrome do membro fantasma (PLS) a un fenómeno de liquidación da columna vertebral. Pacientes con amputación
moitas veces teñen dor de queixo ou formigón na parte do corpo eliminado. Unha posible causa é que as fibras nerviosas no tocón son estimuladas e o cerebro interpreta os sinais como orixinarios da porción amputada. O outro é a reordenación dentro das áreas corticales para que a área diga pola man agora responde a sinais doutras partes do corpo pero aínda os interpreta como chegando á man amputada.
Non obstante, para pacientes sometidos a amputación de extremidades inferiores baixo anestesia epidural, non un dos pacientes 11 que recibiron bloqueo epidural lumbar con bupivacaína e morfina durante 72 horas antes da operación desenvolveu PLS. Para as persoas que sufriron anestesia xeral sen bloqueo epidural anterior, os pacientes 5 de 14 tiveron PLS nas semanas 6 e 3 continuou a experimentar PLS no ano 1.
Woolf e Chong observaron que o tratamento perfecto preoperatorio, intraoperatorio e postoperatorio componse de "AINEs para reducir a activación / centralización dos nociceptores, os anestésicos locais para bloquear o fluxo sensorial e fármacos centralizados como os opiáceos". Aumenta a dor perioperatoria coas técnicas de prevención. A satisfacción, acelerar a descarga, recomenda o uso de opiáceos, xunto con un estreñimiento reducido, sedación, náuseas e retención urinaria e ata pode deter o desenvolvemento da dor crónica. Os anestesiólogos e os cirurxiáns deben considerar a integración destas técnicas nas súas prácticas cotiás.
Cando a dor se produce como consecuencia dun dano ou lesión en consecuencia da cirurxía, a medula espinal pode acadar un estado hiperexcitável no que se producen reaccións de dor excesivas que poden persistir durante días, semanas ou incluso anos.
Por que a lesión localizada resultante do trauma resulta en dor crónica e intratável nalgúns pacientes? A lesión por tecido leva a unha constelación de cambios na excitabilidade espinal, incluíndo un disparo espontáneo elevado, maior amplitude e lonxitude de resposta, umbilidade diminuída, aumento da descarga a repetición de estimulación e campos receptivos expandidos. A persistencia destes cambios, denominados colectivamente sensibilización central, parece ser fundamental para o aumento prolongado da sensibilidade á dor que define a dor crónica. Numerosos fármacos e / ou medicamentos, así como o bloqueo nervioso anestésico local, poden limitar a magnitude do windup do sistema nervioso central (CNS), como se evidencia por diminúe a dor e diminuíu o consumo de opiáceos nos modelos analxésicos preemptivos.
Insight do Dr. Alex Jimenez
O coidado quiropraxia é unha opción de tratamento alternativo que utiliza axustes espinales e manipulacións manuais para restaurar con seguridade e eficacia e manter o correcto aliñamento da columna vertebral. Os estudos de investigación determinaron que os malignos espiñales ou subluxacións poden provocar dor crónica. O coidado quiropraxia utilízase habitualmente para a xestión da dor, mesmo se os síntomas non están asociados a unha lesión e / ou condición no sistema músculo-esquelético e nervioso. Mediante unha alineación da columna vertebral coidadosamente, a Quiroprácticos pode axudar a reducir o estrés e a presión das estruturas que rodean o compoñente principal da fundación do corpo e, finalmente, proporcionar alivio da dor.
Función e dor do sistema nervioso entérico
Cando se trata dun uso reducido de medicamentos e / ou medicamentos, incluídos os opiáceos, a fin de evitar efectos secundarios como problemas de saúde gastrointestinal, a función adecuada do sistema nervioso entérico pode estar en xogo.
O sistema nervioso entérico (ENS) ou sistema nervioso intrínseco é unha das ramas clave do sistema nervioso autónomo (ANS) e consiste nun sistema de nervios en forma de malla que modula o papel do tracto gastrointestinal. É capaz de actuar independentemente do sistema nervioso simpático e parasimpático, aínda que poida verse afectado por eles. O ENS tamén se pode chamar segundo cerebro. Deriva das células da crista neural.
O sistema nervioso entérico en humanos componse dalgunhas neuronas 500 millóns, incluíndo os numerosos tipos de células Dogiel, aproximadamente un dos centésimas da cantidade de neuronas no cerebro. O sistema nervioso entérico insírese no revestimento do sistema gastrointestinal, que comeza no esôfago e se estende ata o ano. As células de Dogiel, tamén coñecidas como células de Dogiel, refírense a algún tipo de tecido adrenal multipolar dentro dos ganglios simpáticos prevertebrales.
O ENS é capaz de funcións autónomas, como a coordinación de reflexos; aínda que recibe unha inervación considerable no sistema nervioso autónomo, funciona e pode funcionar independentemente do cerebro e da medula espiñal. O sistema nervioso entérico foi descrito como o "segundo cerebro" por varias razóns. O sistema nervioso entérico pode funcionar de forma autónoma. Comunícase normalmente co sistema nervioso central (SNC) a través do parasimpático, ou a través do nervio vago, e do simpático, que é a través dos ganglios prevertebrais, o sistema nervioso. Non obstante, estudos de vertebrados revelan que cando se corta o nervio vago, o sistema nervioso entérico segue funcionando.
Nos vertebrados, o sistema nervioso entérico inclúe neuronas eferentes, neuronas aferentes e interneuronas, que fan que o sistema nervioso entérico poida levar reflexos e actuar como centro integrador na ausencia de entrada de SNC. As neuronas sensoriais informan sobre as condicións mecánicas e químicas. O sistema nervioso entérico ten a capacidade de cambiar a súa resposta en función de factores como o nutriente ea composición a granel. Ademais, a ENS contén celas de apoio que son moi parecidas á astroglia do cerebro e unha barreira de difusión en torno aos capilares que rodean os ganglios que son como a barreira hematoencefálica dos vasos sanguíneos.
O sistema nervioso entérico (ENS) xoga un papel fundamental nos procesos inflamatorios e nociceptivos. Os medicamentos e / ou medicamentos que interactúan co ENS suscitaron recentemente un interese considerable debido á súa capacidade para regular numerosos aspectos da fisioloxía e fisiopatoloxía intestinal. En particular, os experimentos en animais demostraron que os receptores activados pola proteína (PAR) poden ser esenciais para a inflamación neuroxénica no intestino. Ademais, os agonistas de PAR2 parecen inducir hipersensibilidade intestinal e estados hiperalxésicos, o que suxire un papel para este receptor na percepción da dor visceral.
Ademais, os PARs, xunto coas proteínas que os activan, representan novos e interesantes obxectivos para a intervención terapéutica na ENS. O alcance da nosa información limítase á quiropráctica e ás lesións e as condicións da columna vertebral. Para discutir o tema, non dubide en preguntar ao Dr. Jimenez ou póñase en contacto connosco 915-850-0900 .
Comisariado polo Dr. Alex Jiménez
Temas adicionais: ciática
Sciática é médicamente referida como unha colección de síntomas, en vez de unha soa lesión e / ou condición. Os síntomas da dor de nervio ciático ou ciática poden variar en frecuencia e intensidade; con todo, descríbese máis comúnmente como unha dor súbita, afiada (afiada) ou eléctrica que irradia desde a parte inferior das costas nas nádegas, cadros, coxas e pernas no pé. Outros síntomas da ciática poden incluír sensacións de formigamento ou queimaduras, entorpecemento e debilidade ao longo do nervio ciático. A ciática afecta a maioría das persoas entre as idades de 30 e 50 anos. A miúdo pode desenvolverse como consecuencia da dexeneración da columna vertebral debido á idade, con todo, a compresión e irritación do nervio ciático causada por un abultamiento ou disco herniado, entre outras cuestións de saúde espiñal, tamén pode causar dor de nervio ciático.
A ferramenta Find A Practitioner de IFM é a rede de referencia máis grande en Medicina Funcional, creada para axudar aos pacientes a localizar a profesionais da Medicina Funcional en calquera parte do mundo. Os profesionais certificados IFM aparecen en primeiro lugar nos resultados da busca, dada a súa ampla educación en Medicina Funcional

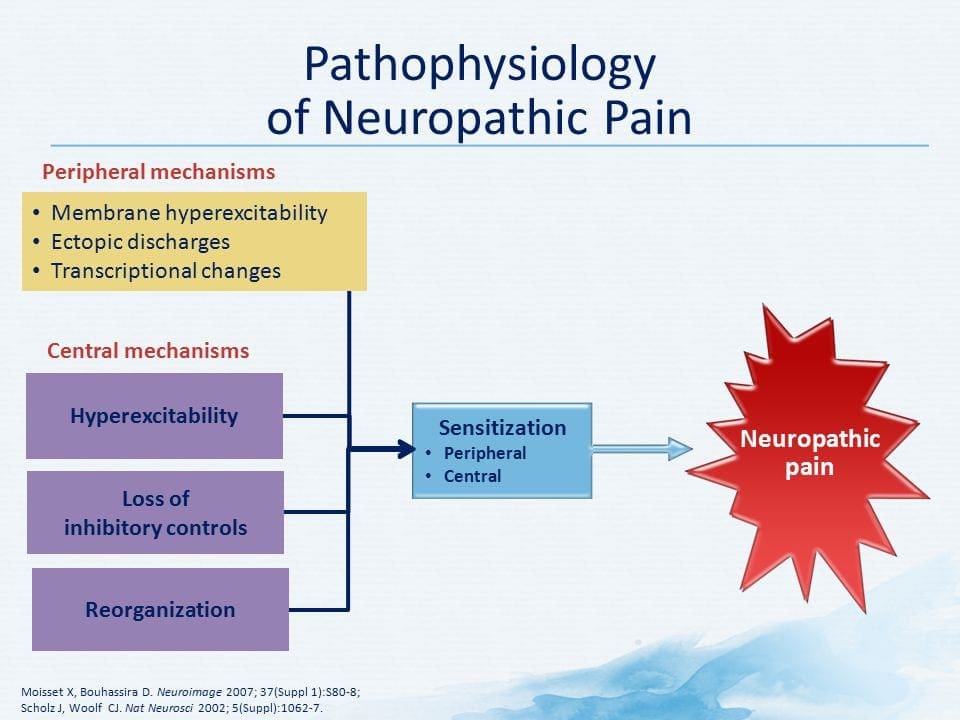
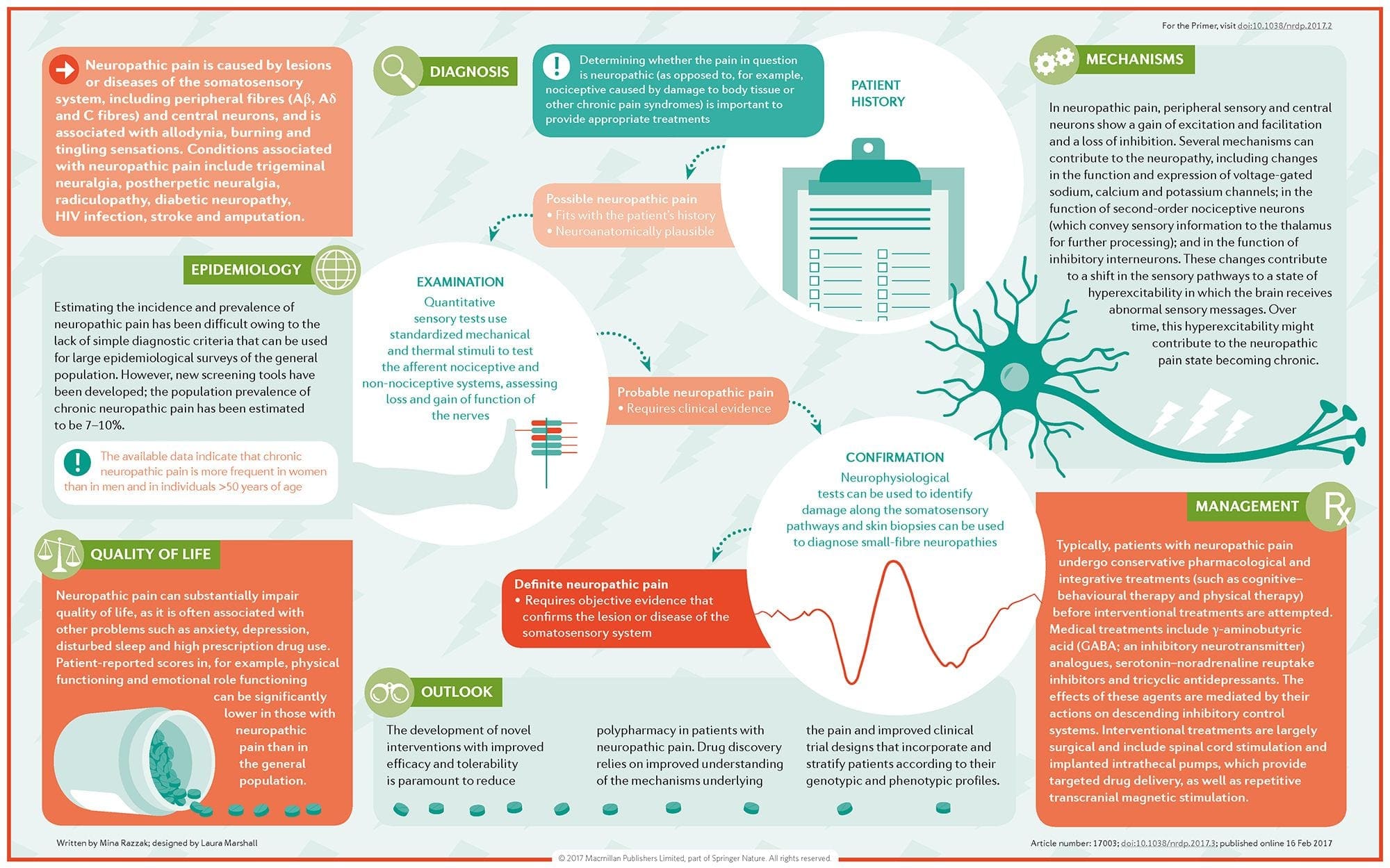




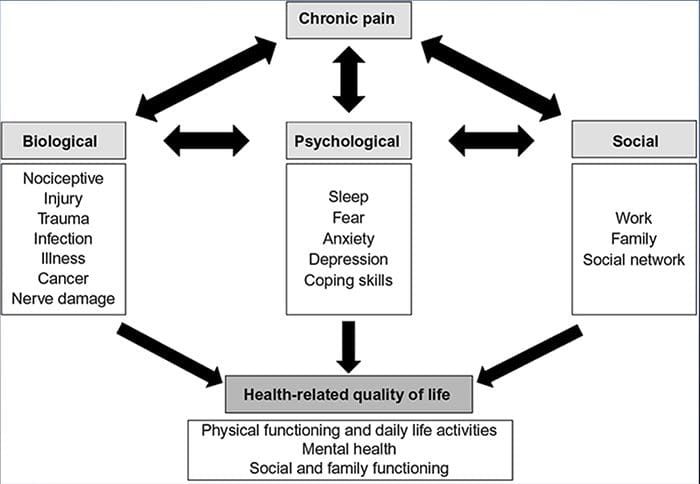
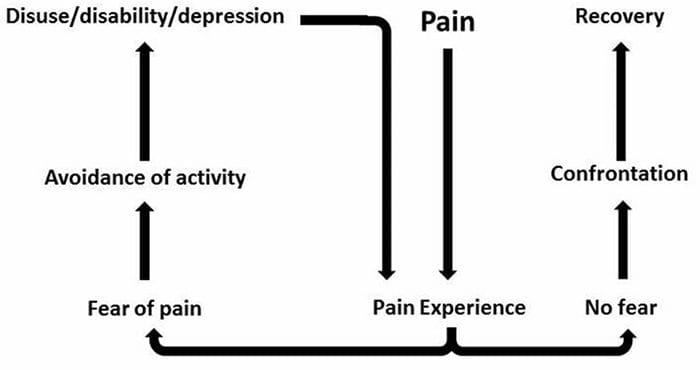
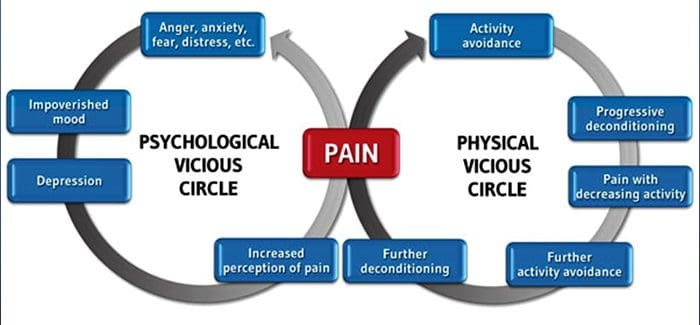
















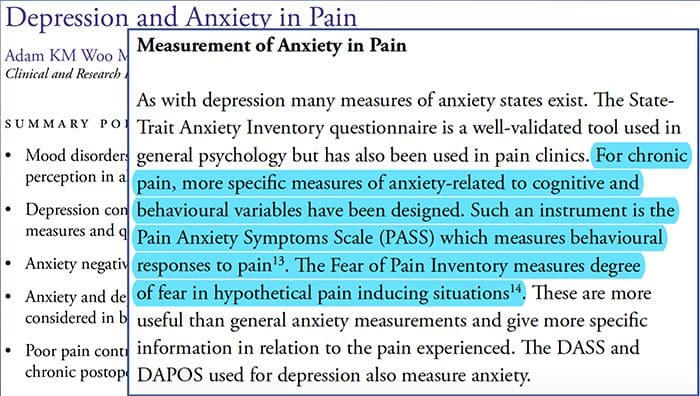


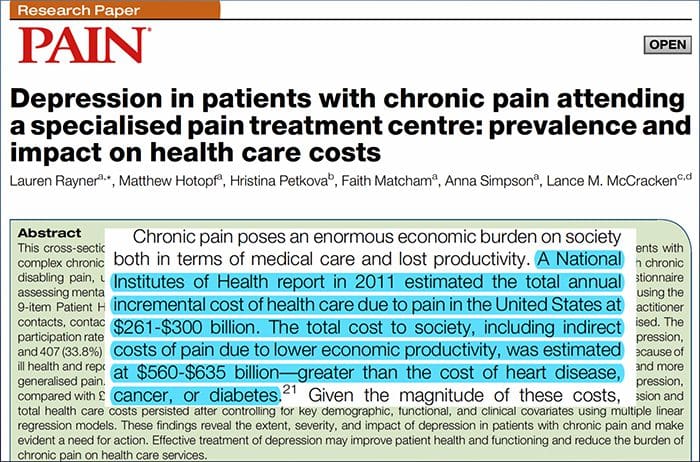



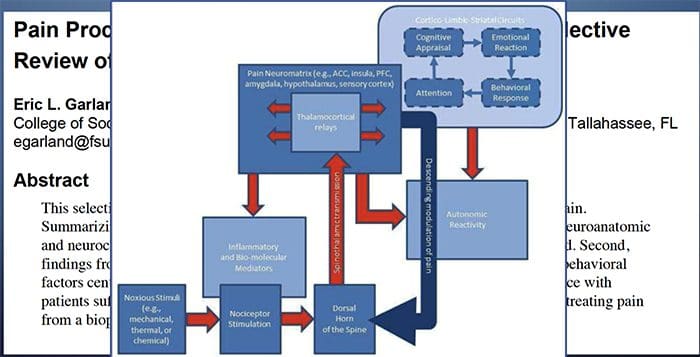






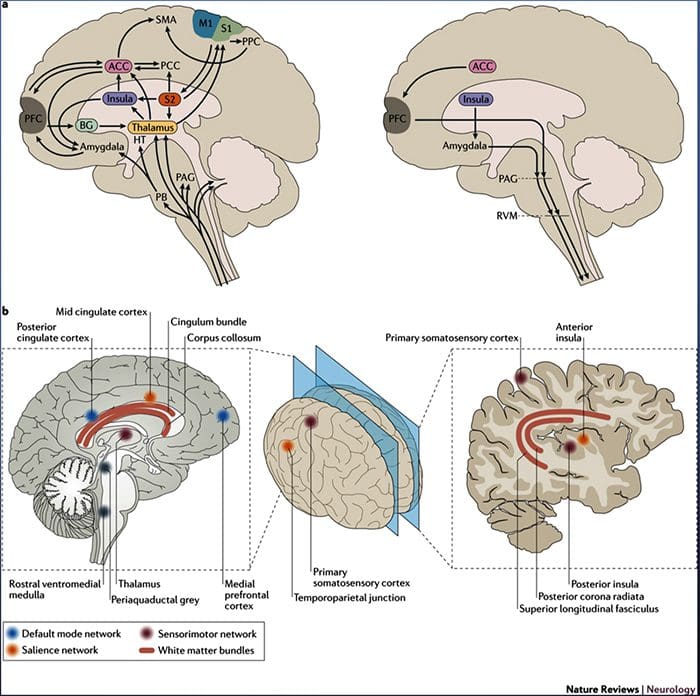

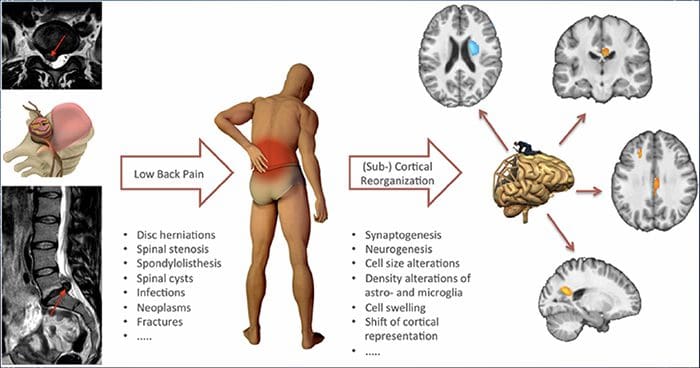

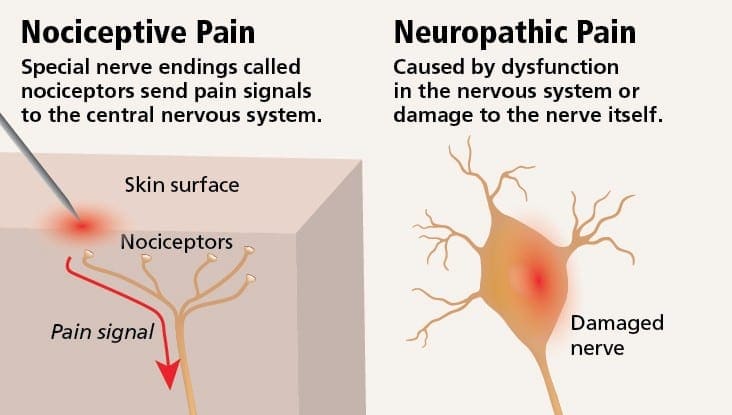
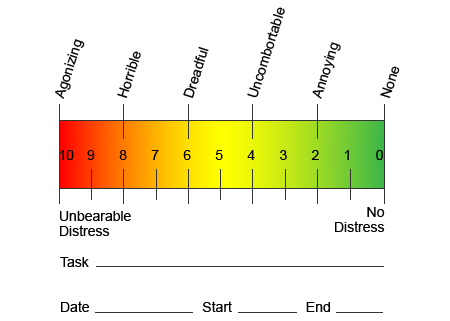


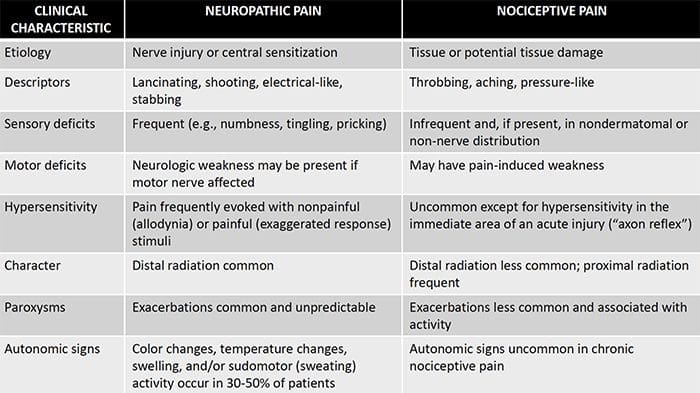
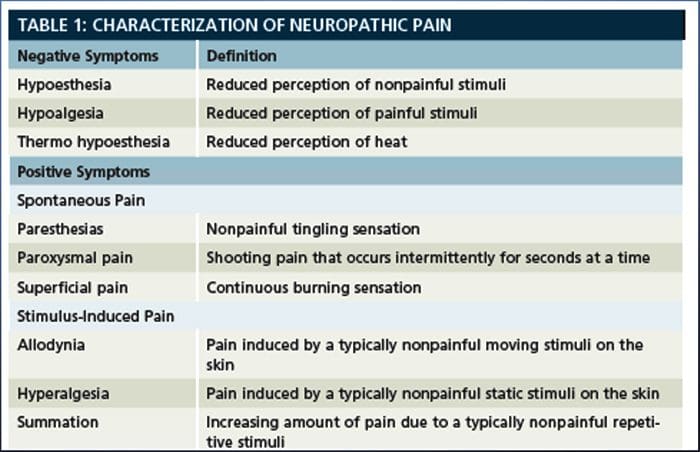
 PATOGENESIS DO DOLOR NEUROPÁTICO
PATOGENESIS DO DOLOR NEUROPÁTICO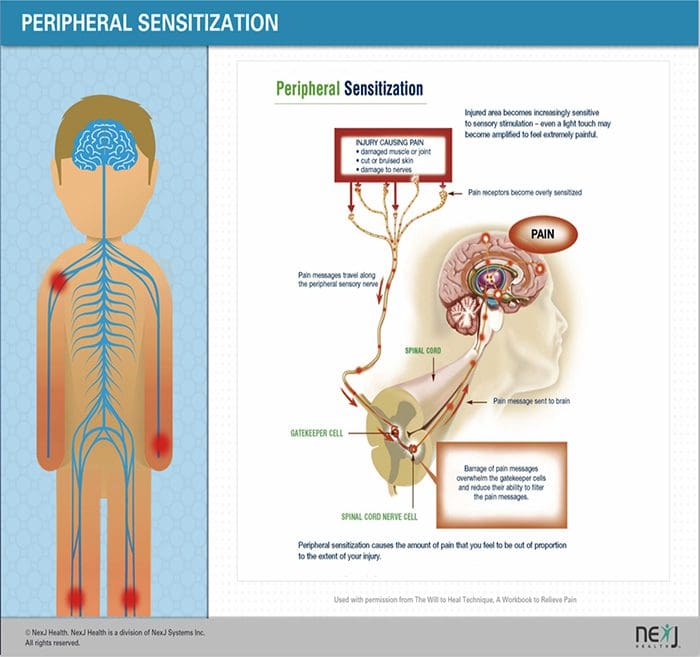
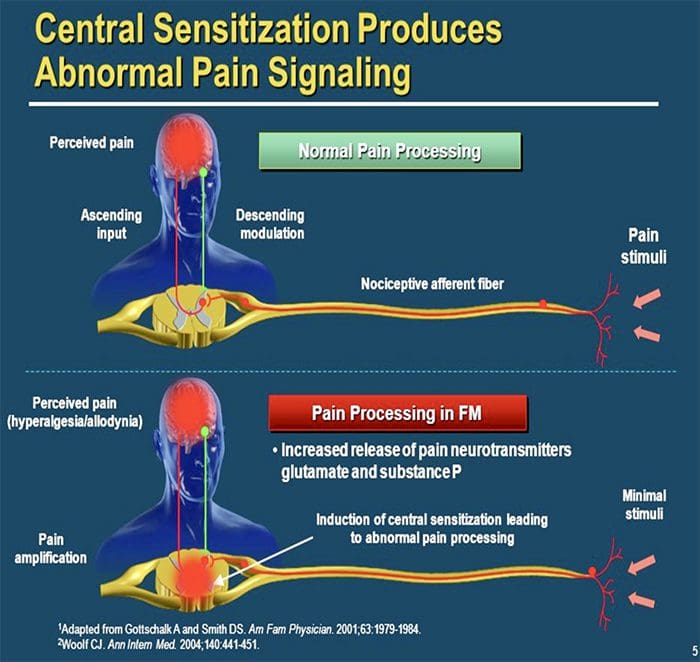
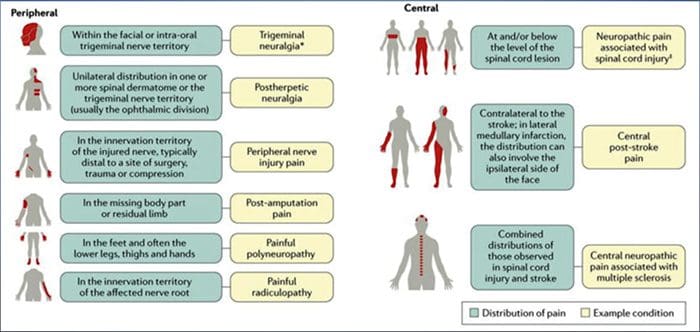 CAUSAS COMUNES
CAUSAS COMUNES
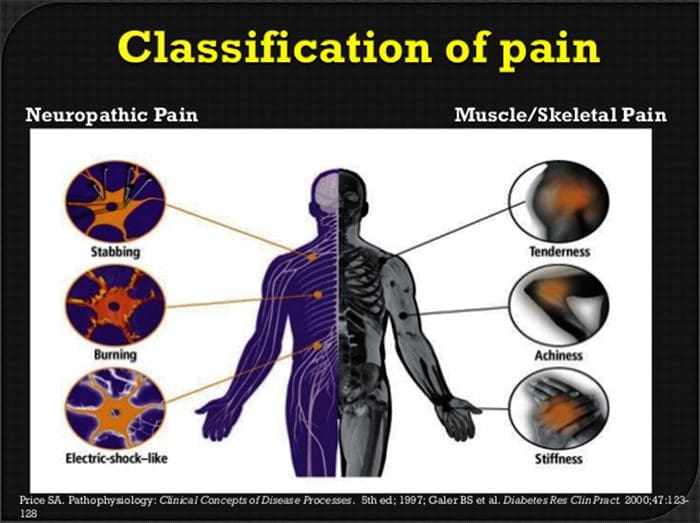
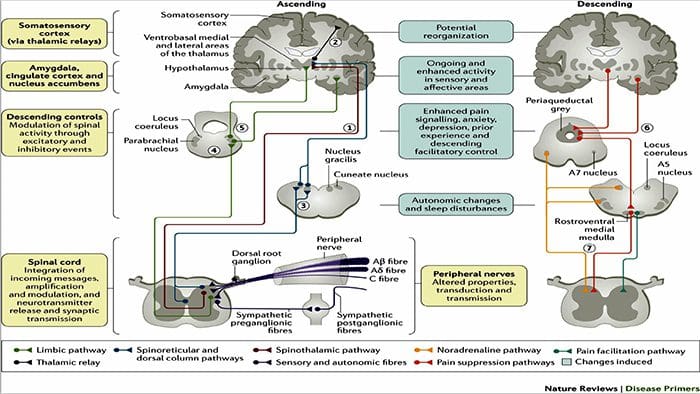
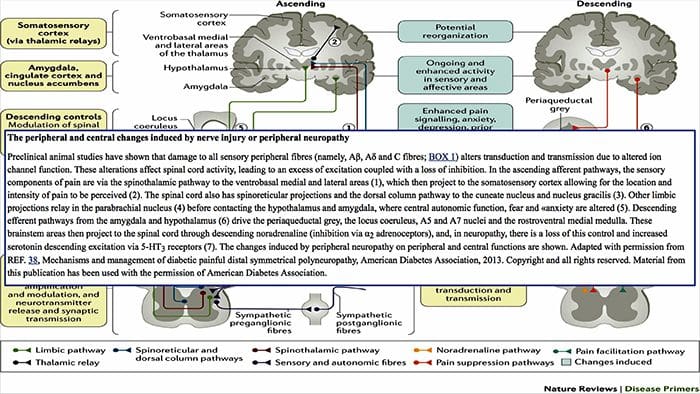

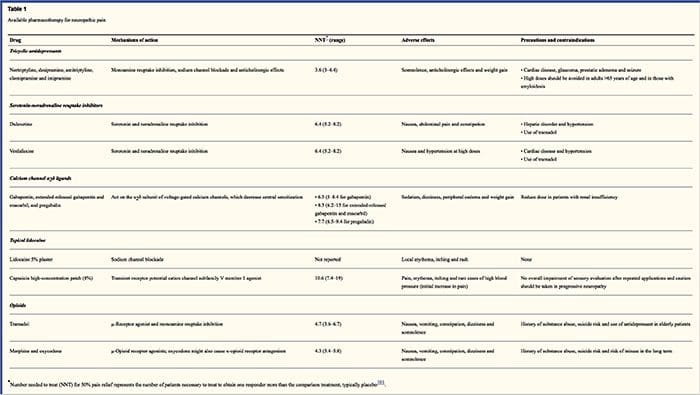




 DOR DE EXTREMIDADE FANTÁMICA E REALIDADE AUGMENTADA
DOR DE EXTREMIDADE FANTÁMICA E REALIDADE AUGMENTADA
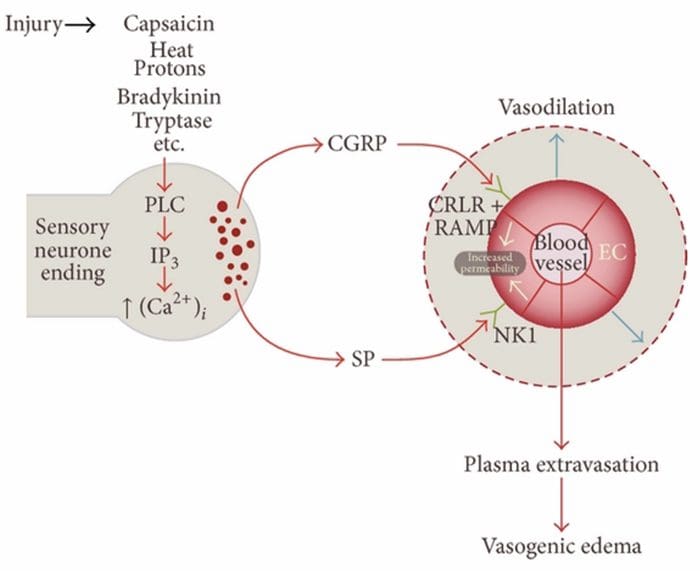 CAUSAS COMUNES
CAUSAS COMUNES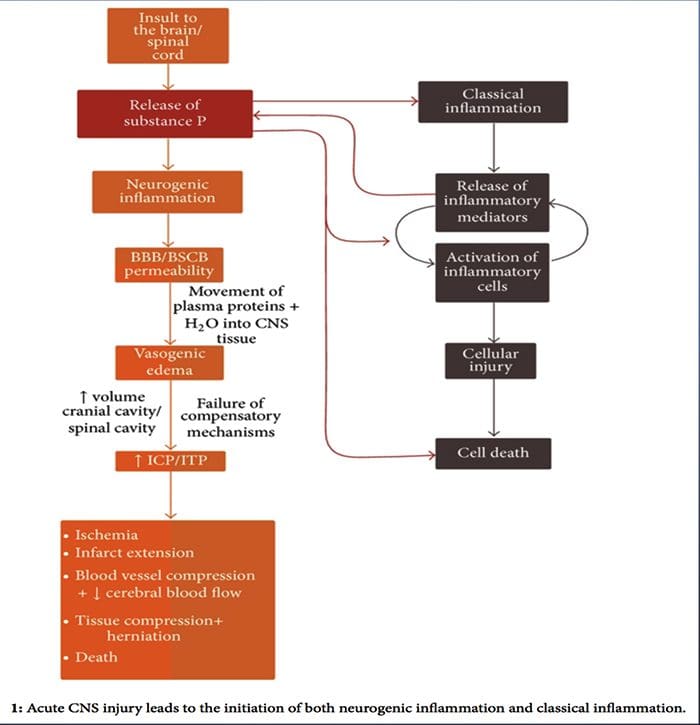


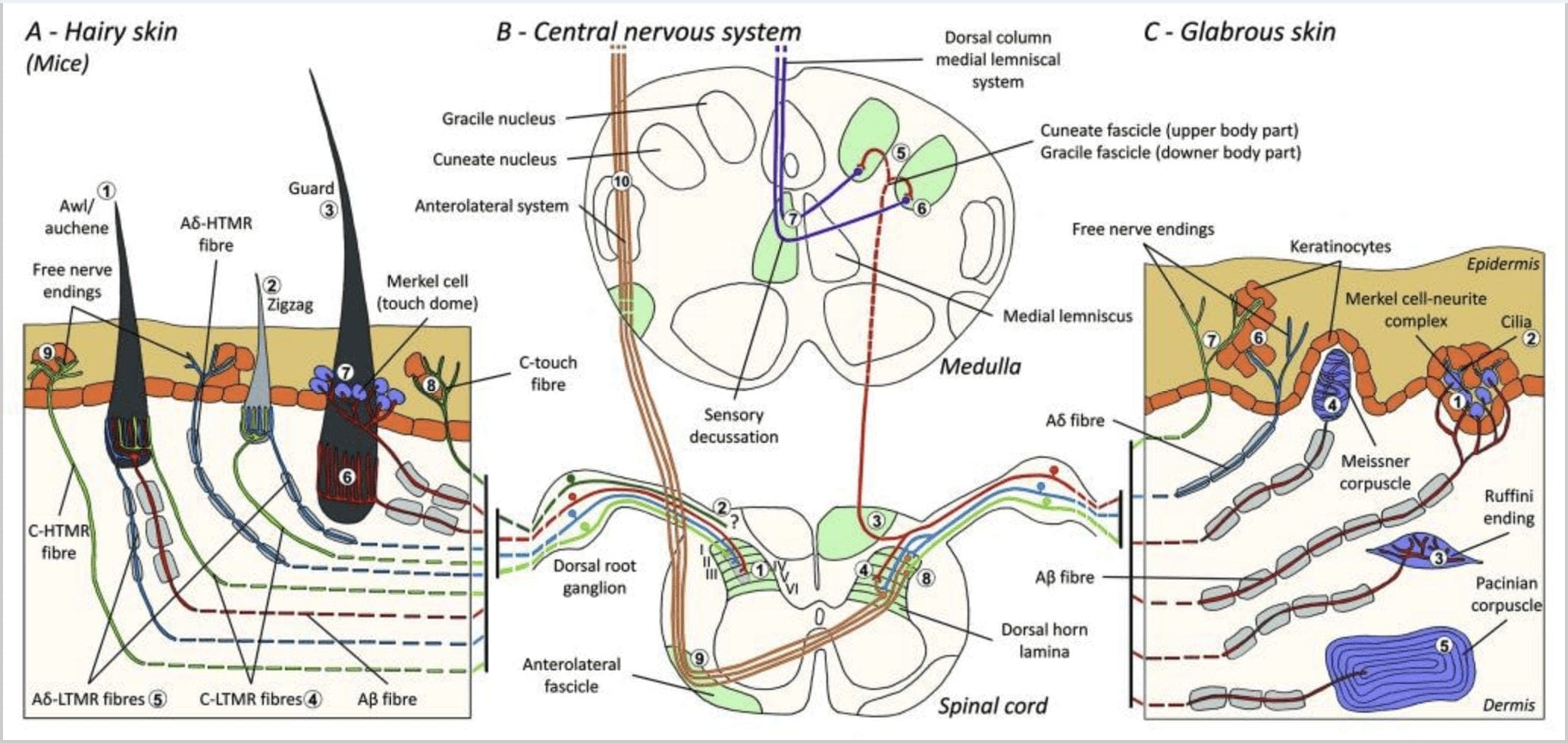
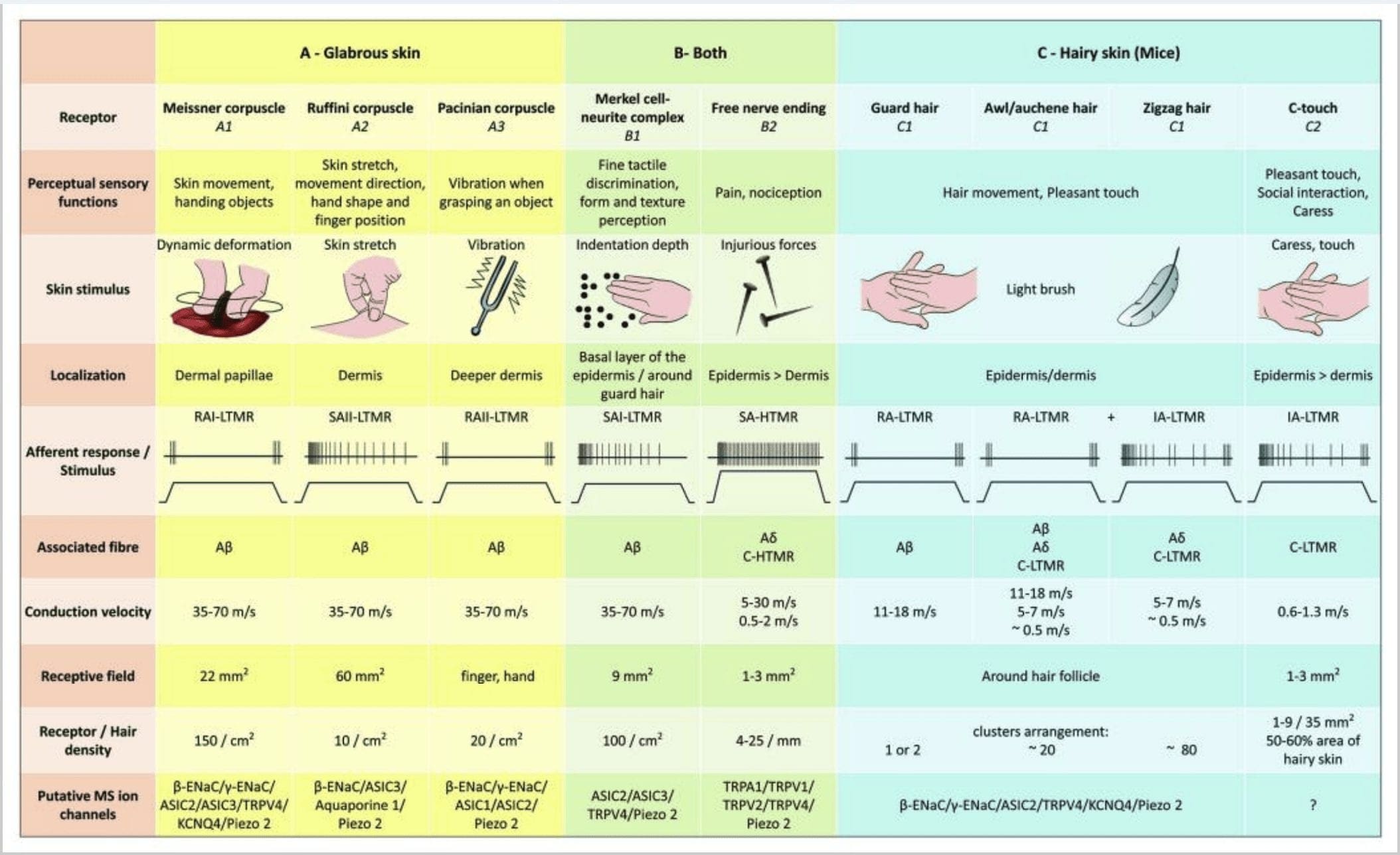

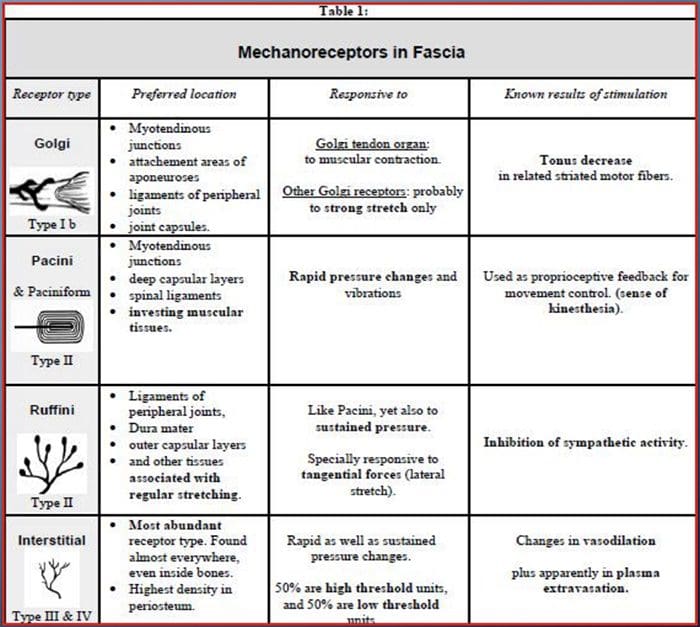 TEORÍA DO CONTROL DA PORTA DA DOR
TEORÍA DO CONTROL DA PORTA DA DOR
 COLUMNA DORSAL VÍA LEMNISCAL MEDIAL
COLUMNA DORSAL VÍA LEMNISCAL MEDIAL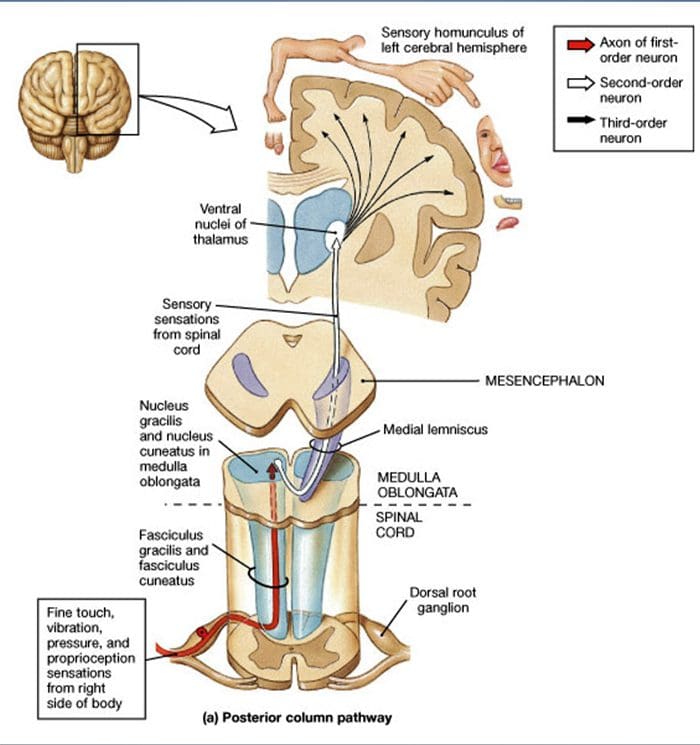 UTILIZACIÓN DE MECANORECEPTORES PERIFÉRICOS PARA ALTERAR A PERCEPCIÓN DA DOR
UTILIZACIÓN DE MECANORECEPTORES PERIFÉRICOS PARA ALTERAR A PERCEPCIÓN DA DOR
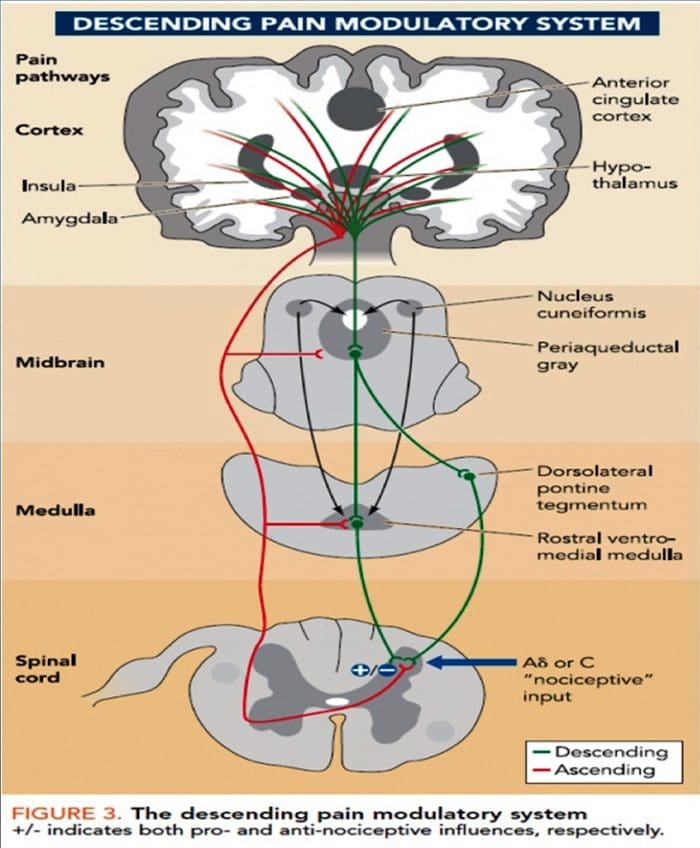
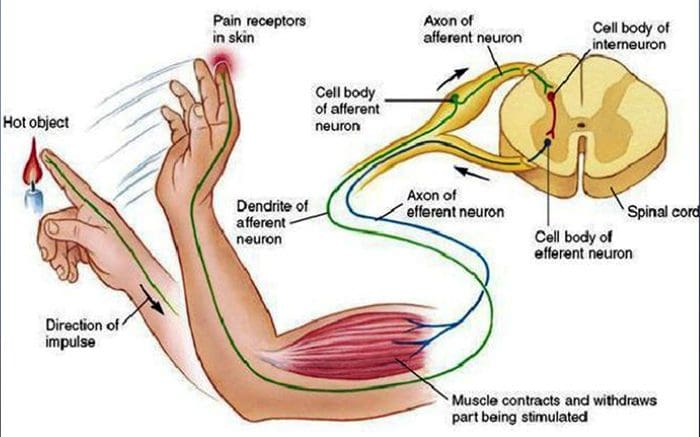
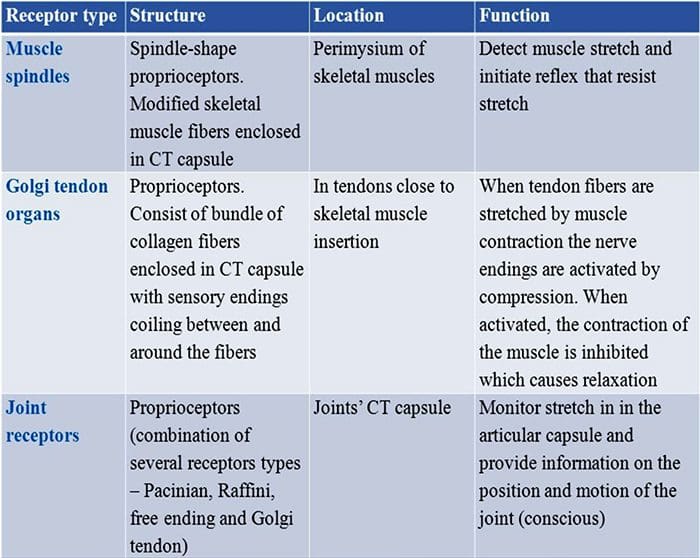
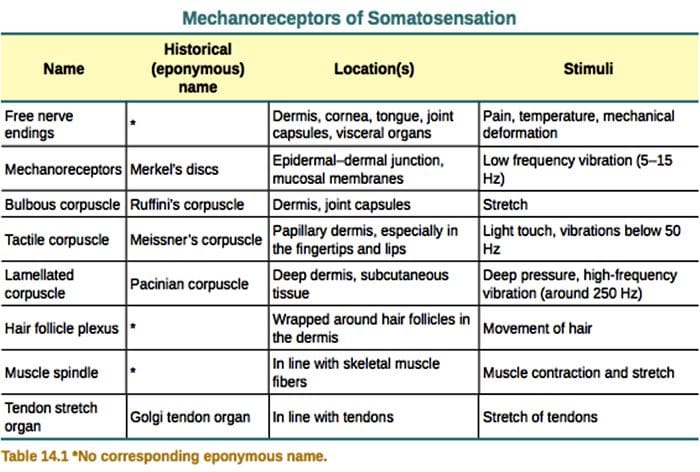 Vibrado
Vibrado
 TOCO DE LUZ
TOCO DE LUZ



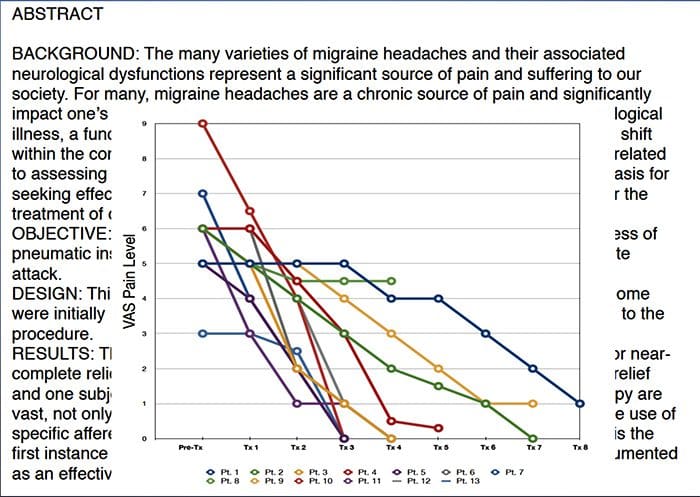


 ESTUDO DE CASO
ESTUDO DE CASO DESTACADOS DE EXAMENO FÍSICO
DESTACADOS DE EXAMENO FÍSICO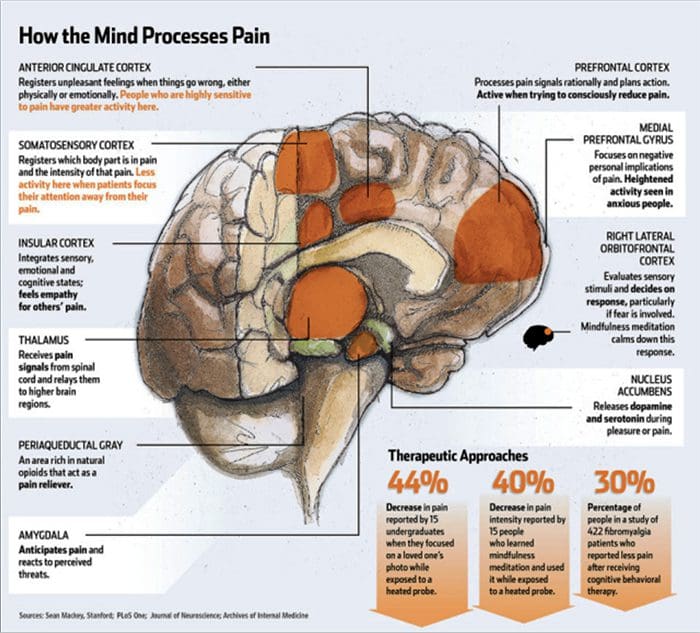 DESTACADOS DE EXAMENO FÍSICO
DESTACADOS DE EXAMENO FÍSICO
 DESPOIS DE DOUS DÍAS
DESPOIS DE DOUS DÍAS